FEDERAL COURT OF AUSTRALIA
Commonwealth Scientific and Industrial Research Organisation v BASF Plant Science GmbH [2020] FCA 328
ORDERS
COMMONWEALTH SCIENTIFIC AND INDUSTRIAL RESEARCH ORGANISATION Appellant | ||
AND: | Respondent | |
DATE OF ORDER: |
THE COURT ORDERS THAT:
2. The decision of the Commissioner of Patents made on 10 December 2018 be set aside.
3. Within 21 days of the date hereof the parties file and serve proposed minutes of orders and short submissions (limited to 3 pages) as to any necessary consequential orders including on the question of costs.
Note: Entry of orders is dealt with in Rule 39.32 of the Federal Court Rules 2011.
BEACH J:
1 The Commonwealth Scientific and Industrial Research Organisation (CSIRO) appeals from a decision of a delegate of the Commissioner of Patents allowing amendments to Australian patent application 2013273704 entitled “Process for the production of polyunsaturated fatty acids in transgenic organisms” (the application) filed by BASF Plant Science GmbH (BASF).
2 The application relates to genes from a species of unicellular algae that code for enzymes which can be employed for the recombinant production of polyunsaturated fatty acids in plants. Broadly, it is directed to the production of long-chain polyunsaturated fatty acids in higher plants by incorporating genes that encode for relevant enzymes from microalgae, specifically one called Ostreococcus lucimarinus.
3 CSIRO contends that the amendments are not allowable because, as a result of the amendments, the amended specification would claim and disclose matter that extends beyond that disclosed in the complete specification as filed, and therefore are impermissible under s 102(1) of the Patents Act 1990 (Cth) (the Act).
4 The application is a divisional application of Australian patent application 2007304229 filed on 4 October 2007. But the application claims priority from European patent application 06121888.9 filed on 6 October 2006 (the priority date).
5 The application was filed on 19 December 2013. BASF requested three amendments to the application as filed prior to acceptance; those amendments were requested on 7 September 2015, 26 February 2016 and 26 April 2016.
6 The application was accepted with those amendments on 4 May 2016. The application as accepted was published on 19 May 2016. CSIRO filed a notice of opposition to the application as accepted on 19 August 2016.
7 On 7 June 2017, BASF filed a request to amend the specification of the application as accepted under s 104. That request was not allowed on the basis that, as a result of the proposed amendment, the specification would not comply with the requirements of s 40(3).
8 On 2 October 2017, BASF filed a further request to amend the specification of the application as accepted.
9 Details of these two requests were published in the Australian Official Journal of Patents on 30 November 2017. CSIRO filed a notice of opposition to any allowance of those amendments on 30 January 2018.
10 On 10 December 2018, a delegate of the Commissioner of Patents allowed the post-acceptance amendments sought. CSIRO has appealed that decision under s 104(7). The nature of the appeal before me is by way of a rehearing de-novo.
11 Now whether the proposed post-acceptance amendments are allowable is a question to be determined under s 102(1) of the Act as amended by the Intellectual Property Laws Amendment (Raising the Bar) Act 2012 (Cth) (the Raising the Bar Act). The amendments made to s 102(1) of the Act by item 29 of Sch 1 of the Raising the Bar Act apply in relation to amendments of complete specifications directed or requested to be made on or after 15 April 2013 if the amendments are in relation to complete patent applications made on or after that day (Raising the Bar Act, sch 1, item 55(9)(d)). As I have said, the application was filed on 19 December 2013.
12 Section 102(1) in its present and applicable form provides:
(1) An amendment of a complete specification is not allowable if, as a result of the amendment, the specification would claim or disclose matter that extends beyond that disclosed in the following documents taken together:
(a) the complete specification as filed;
(b) other prescribed documents (if any).
13 For present purposes, the only “other prescribed document” (reg 10.2A of the Patents Regulations 1991 (Cth)) is the abstract that accompanied the application. But this is identical to page 1, lines 7 to 13 of the application as filed. Accordingly, consideration of the abstract adds nothing to the analysis.
14 Now it is not in doubt that the matter arising “as a result of the amendment” is identified by identifying the difference between the application as accepted and the application as proposed to be amended. One then compares what is disclosed in the application as proposed to be amended, of course read as a whole, with what is disclosed in the application as filed. If as a result of the amendment proposed, the specification would disclose or claim matter that extends beyond that disclosed in the application as filed, the amendment is not allowable. Moreover, part of the analysis concerning construing the application as filed is to be undertaken through the eyes of a person skilled in the art at the relevant time.
15 On the present appeal CSIRO filed and served expert evidence from Dr Surinder Singh who was at the time of preparing his written evidence a Group Leader and Chief Research Scientist in CSIRO’s Agriculture and Food Unit. BASF filed and served expert evidence from Dr David Stalker. The experts met in conclave and subsequently produced a joint expert report. I took their evidence in a concurrent evidence session with the debate between them largely focused upon the question of substrate specificity concerning polyunsaturated fatty acids that was addressed in the application as filed.
16 As I have indicated, the application relates to the field of isolating polynucleotides which code for enzymes that can be used in the biosynthetic pathway of long-chain polyunsaturated fatty acid synthesis and introducing these polynucleotides into plants (e.g. oil seed crops) by genetic engineering with the intention that long-chain polyunsaturated fatty acids will be produced in such plants. The work in this field is predominantly done by senior research scientists with significant research experience in relation to the synthesis of fatty acids in transgenic plants. In the present case, the hypothetical person skilled in the art at the relevant time is such a scientist involved in metabolic/genetic engineering of plants to produce fatty acids.
17 Dr Singh has a PhD from the University of Adelaide in plant physiology and biochemistry. He has extensive experience in metabolic/genetic engineering of plants for the development of plant oils, including fatty acids. He is a named author of 102 peer-reviewed articles and papers, including 75 publications directed to plant lipids and modifying plant oils. He is a person skilled in the field of the application.
18 Dr Stalker also holds a PhD from the University of Cincinnati in the field of cell biology and bio-chemistry. He has worked as a scientist in the field of molecular biology for over 45 years. For a substantial part of that period he has researched and commercialised innovations in plant biotechnology, including creating genetically engineered plants. He has also published a substantial number of peer-reviewed articles and papers in this field. He has been a professor of biotechnology at RMIT and is the founder and a director of Generic Genetics Group Pty. Ltd. He is similarly skilled in the relevant field.
19 For the reasons that follow, I am clearly satisfied that the proposed amendments are impermissible and that the appeal should be allowed.
20 Let me begin with some science before turning to discuss and compare the application as filed, the application as accepted and the application as proposed to be amended.
Some Science
21 Let me begin by setting out a functional glossary, that is, functional to following my reasons and functional to appreciating the context of the invention the subject of the application, of the following terms:
Alpha(α)-linolenic acid
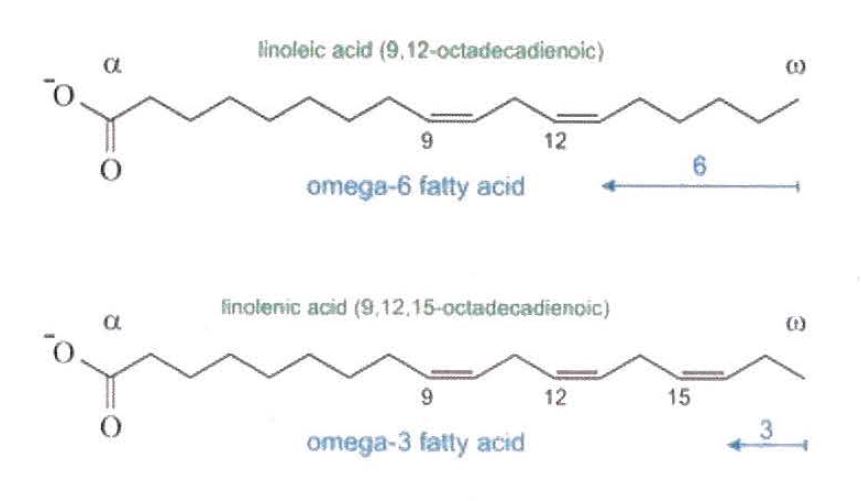
The starting material for the synthesis of omega(ω)-3 long-chain polyunsaturated fatty acids.
CoA-dependent enzyme
An enzyme that predominantly acts on acyl-Coenzyme A (CoA) bound fatty acid substrates in the cytosolic acyl-CoA pool as opposed to acting on phosphatidylcholine bound fatty acid substrates in the endoplasmic reticulum-associated phospholipid pool.
Desaturase
An enzyme that catalyses the insertion of a carbon to carbon double bond in the carbon backbone by removing two hydrogen atoms from a fatty acid.
Elongase
An enzyme that catalyses the addition of two carbon atoms to the terminal carboxyl end of the fatty acid, extending the length of a fatty acid.
Enzyme
A protein molecule that functions as a catalyst for biochemical reactions.
Fatty acid
A carboxylic acid consisting of a chain of carbon atoms bonded to hydrogen atoms (forming a carbon backbone) having a carboxyl group (COOH) at one end and a methyl group (CH3) at the other end.
Linoleic acid
The starting material for the synthesis of omega(ω)-6 long-chain polyunsaturated fatty acids, for example arachidonic acid.
Long chain PUFA (LC-PUFA)
A fatty acid having a carbon backbone that is at least 20 carbon atoms long and contains two or more double bonds.
Ostreococcus lucimarinus (O. lucimarinus)
A unicellular algae.
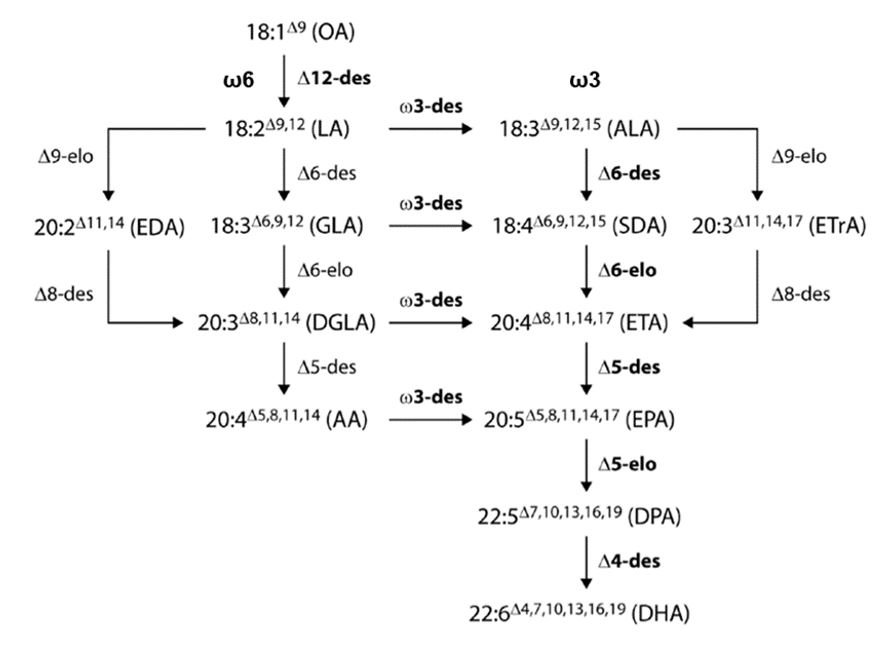
Omega(ω)-3 fatty acid
A fatty acid with the last carbon to carbon double bond three carbon atoms away from the terminal methyl group, for example, eicosapentaenoic acid and docosahexaenoic acid.
Omega(ω)-6 fatty acid
A fatty acid with the last carbon to carbon double bond six carbons atoms away from the terminal methyl group.
Polynucleotide
A nucleic acid sequence made up of nucleotides that code for genetic information.
Polypeptide
A sequence of amino acids forming a protein.
Polyunsaturated fatty acids (PUFAs)
Fatty acids having two or more carbon to carbon double bonds in the fatty acid chain. PUFAs are categorised by the number of carbon atoms in the chain, the number of unsaturated (double) bonds, and the position of those double bonds. PUFAs can be classified into two main groups, namely, omega(ω)-3 fatty acids and omega(ω)-6 fatty acids.
Recombinant production/recombinant methods
A combination of genetic material (DNA) from different organisms.
Saturated fatty acid
A fatty acid containing no carbon to carbon double bonds.
Specificity
For the moment I will define this to be the ability of an enzyme to discriminate between competing potential substrates.
Substrate
The molecule upon which an enzyme acts.
Unsaturated fatty acid
A fatty acid having at least one carbon to carbon double bond, rather than only single bonds.
22 It is also necessary to set out the following list of the fatty acids referenced in the relevant biosynthetic pathway that I will discuss later:
List of fatty acids referenced in the biosynthetic pathway
Abbreviation | Carbons (No of carbons: no of double bonds) | Name |
OA | 18:1 | Oleic acid |
LA | 18:2 | Linoleic acid |
ALA | 18:3 | Alpha-linolenic acid |
GLA | 18:3 | Gamma-linolenic acid |
SDA | 18:4 | Stearidonic acid |
EDA | 20:2 | Eicosadienoic acid |
ETrA | 20:3 | Eicosatrienoic acid |
DGLA | 20:3 | Dihomo-gamma-linolenic acid |
ETA | 20:4 | Eicosatetraenoic acid |
AA (or ARA) | 20:4 | Arachidonic acid |
EPA | 20:5 | Eicosapentaenoic acid |
DPA | 22:5 | Docosapentaenoic acid |
DHA | 22:6 | Docosahexaenoic acid |
(a) Enzymes
23 Let me begin my discussion of some of the science by talking about enzyme function. What I have set out below on this topic and later biochemical topics is a composite of the evidence from the two experts that I did not understand to be in issue concerning the applicable common general knowledge relevant to the task of construing the application as filed, accepting of course that the relevant legal questions are for me as to what it discloses.
24 In a biological setting, enzymes are typically proteins which function as catalysts for biochemical reactions. An enzyme works on a molecule known as the substrate, and the molecules formed by the reaction are known as the products.
25 A protein is made up of one or more chains of amino acids. A protein will be an enzyme if it performs the role of catalysing a biochemical reaction. The chemical structure that a protein enzyme has and its function is dictated by the particular order or sequence of the amino acids that make up that enzyme. Changing that sequence of amino acids may create a different protein enzyme, which may or may not then have a different function.
26 The sequence of amino acids that form a protein enzyme is in turn dictated by the gene sequence which encodes the protein enzyme. A gene sequence is a chain of nucleotide bases. The information in a gene sequence is stored as a “code” utilising four nucleotides, albeit in triplets of three nucleotides known as codons. The four nucleotides are adenine (A), cytosine (C), guanine (G) and thymine (T). The sequence of these nucleotides determines the information available for building and maintaining an organism. I have discussed the relevant concepts elsewhere (Meat and Livestock Australia Ltd v Cargill Inc (2018) 354 ALR 95 (Meat and Livestock (No 1)) at [21] to [66]). Judicial notice can be taken of the fact that such concepts have been well known to biochemists, biologists and experts in derivative fields for some decades.
27 Whilst a particular gene sequence may encode a specific protein enzyme, it is possible that a different gene sequence may still encode the same enzyme. So, the same protein enzyme could be encoded by two or more different gene sequences. Different codons can code for the same amino acid, for example, TTT and TTC both code for phenylalanine. So, variation at the DNA level does not necessarily produce a protein having altered structure and function.
28 On the other hand, even a single nucleotide change to a DNA sequence that results in a single amino acid change at a given point can change the structure of the encoded protein enzyme and therefore alter its function. For example, a single nucleotide change such as TTT to TAT produces a different amino acid that has different biochemical properties; in this example the change is from phenylalanine (non-polar) to tyrosine (polar).
29 Now an enzyme works on the substrate. An enzyme may bind to and convert a single substrate only. Alternatively, an enzyme may bind to and convert multiple substrates. An enzyme may also catalyse more than one reaction. For example, an enzyme may only act on molecules that have specific functional groups, for example amino, phosphate or methyl groups, or on a particular type of chemical bond, or on a particular steric or optical isomer.
30 Generally speaking, the “substrate specificity” of an enzyme refers to the ability of that enzyme to discriminate among competing potential substrates and to bind to a specific substrate or substrates. I will return to this issue and discuss it in more detail later. Substrate specificity is the result of structural and conformational complementarity between the enzyme and the relevant substrate or substrates. Where an enzyme can bind to more than one substrate, the enzyme may show a preference for binding to one substrate over another at differing rates or even absolutely.
31 Whether an enzyme has specificity for one or more relevant substrates or is able to effectively perform one or more catalytic reactions will be determined by the amino acid sequence of the enzyme. Consequently, variation in amino acid sequence may influence whether an enzyme might bind to one or multiple substrates or is able to perform one or more catalytic reactions. Sequence variation may also influence an enzyme’s affinity for one or more substrates, therefore impacting substrate specificity for one or more substrates and the enzyme’s preference when able to bind to more than one substrate.
32 Some enzymes can catalyse more than one type of reaction. An enzyme that catalyses two distinct reactions is known as bifunctional. Further, the substrate specificity for the enzyme may depend on the relevant function or catalytic activity. Further, the substrate specificity for one catalytic activity may be different from the substrate specificity for the other catalytic function. In other words, substrate specificity may need to be considered in the context of a relevant function or catalytic activity.
33 The efficiency of the enzyme to convert a substrate or substrates into a relevant product or products refers to the rate at which the substrate is converted into the product. Efficiency can be dependent on amino acid sequence. A change in sequence could alter efficiency by altering substrate specificity, the strength at which the enzyme binds to the substrate being its binding affinity or the rate of catalysis.
34 Enzymes that perform the same or a similar function may have a degree of sequence relatedness, both in terms of the sequence of amino acids that make up each enzyme, and in terms of the DNA sequence that encodes for each enzyme.
(b) Polyunsaturated fatty acids
35 Let me now say something about fatty acids and particularly polyunsaturated fatty acids (PUFAs) and long-chain polyunsaturated fatty acids (LC-PUFAs).
36 A fatty acid is the basic building block of fats and lipids in plants, animals, microorganisms and food. A fatty acid is a carboxylic acid consisting of a hydrocarbon chain that forms a carbon backbone. At one end of the carbon backbone is a carboxyl group (COOH), which is called the terminal carboxyl end, and at the other end of the chain is a methyl group (CH3), which is called the terminal methyl end. Fatty acids are either saturated, that is, the carbon backbone contains no double bonds between the carbon atoms, or unsaturated, that is, the carbon backbone contains one or more carbon to carbon double bonds (C=C double bonds). Monounsaturated fatty acids contain one C=C double bond, and PUFAs are fatty acids that contain more than one C=C double bond.
37 Fatty acids differ in the length of the carbon backbone, and can be categorised from short chain fatty acids to very long chain fatty acids. They contain an even number of carbons in the carbon backbone. LC-PUFAs are fatty acids that contain a carbon backbone that is at least 20 carbons long and contains multiple C=C double bonds.
38 PUFAs can be characterised as either omega-3 (ω3) or omega-6 (ω6) fatty acids based on the position of the first C=C double bond from the terminal methyl group. In particular, ω3 fatty acids have the last C=C double bond 3 carbon atoms away from the terminal methyl group whereas ω6 fatty acids have the last C=C double bond 6 carbon atoms away from the terminal methyl group.
39 LC-PUFAs have important roles in human health and development, with many studies predating the priority date indicating that deficiencies in these fatty acids can increase, for example, the risk or severity of cardiovascular disease, inflammatory diseases, and neuropsychiatric disorders such as depression or dementia.
40 Nutritionally important ω6 LC-PUFAs include arachidonic acid (AA) and nutritionally important ω3 LC-PUFAs include eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). These fatty acids are either directly available as components of the diet or produced from the two essential ω6- and ω3- fatty acids, linoleic acid (LA) and α-linolenic acid (ALA). Although humans can synthesize both EPA and DHA from ALA, direct uptake of EPA and DHA from the diet, for example from marine fish (fish oils), is a significantly more important source of ω3 LC-PUFAs for humans.
41 As a result of a plateau and potentially a decline in marine fish stocks since the early 1990s, significant research effort has focused on producing ω3 LC-PUFAs in alternative sources including aquaculture, cultured microalgae and oilseed crops that have been engineered to produce ω3 LC-PUFAs, such as EPA and DHA. Because of the production capacity and relatively low production cost of PUFAs from plants, oilseed crops that had been genetically engineered to synthesise fatty acids were viewed as a significant goal as a sustainable alternative source of ω3 LC-PUFAs.
42 Before the priority date, transgenic plants had already been produced which synthesized EPA, docosapentaenoic acid (DPA) and DHA in their seed.
43 Prior to the priority date, researchers had been experimenting with introducing foreign genes into plants to alter biological properties and/or to add or enhance physical attributes, such as resistance to stress and disease or herbicide resistance. A plant that is modified using genetic techniques is usually referred to as a genetically engineered plant or a transgenic plant.
44 In a transgenic plant, the source of any introduced foreign gene is relevant insofar as the DNA or amino acid sequence determines the encoded protein’s structure and/or function, such that the introduction of a foreign gene encoding an enzyme from one source can have a different impact in the transgenic plant compared to introduction of a foreign gene encoding an enzyme of a different source, even where both enzymes have the same or similar function.
(c) Synthesising long-chain polyunsaturated fatty acids
45 Let me elaborate further on LC-PUFA plant metabolic engineering.
46 Prior to the priority date research groups were working on isolating and cloning genes from various fungi, protists and microalgae that encode each of the desaturases and elongases involved in the aerobic pathway for LC-PUFA synthesis. The availability of genes that encode the aerobic LC-PUFA biosynthetic enzymes led to attempts to introduce these genes into plants (such as oil seed crops) via recombinant methods and to express these enzymes in these plants to produce LC-PUFAs. Recombinant methods of course combine DNA sequences from different organisms, creating a new recombinant DNA molecule that is not otherwise found in nature.
47 As I have indicated, fatty acids form an important component of soluble fats in living cells and consist of a chain of an even number of carbon atoms, with hydrogen atoms along the length of the chain. At one end of the chain is a methyl group. A carboxyl group is at the other end.
48 And as I have said, PUFA is a term used to describe fatty acid chains that contain two or more carbon-carbon double bonds. In plants, PUFAs occur predominantly as chains having 16, 18, 20 or 22 carbons, hence are referred to as C16-, C18-, C20- and C22-PUFAs, respectively.
49 LC-PUFAs (20 carbons or longer) such as EPA and DHA play important roles in human health and are intrinsic to diet.
50 As I have said, humans can synthesise LC-PUFAs through dietary intake of precursor fatty acids. Humans can also obtain LC-PUFAs via direct dietary intake, e.g. by consuming fish and fish-derived oils that contain EPA and DHA. However, poor diets combined with decreasing global fish stocks have raised the importance of obtaining LC-PUFAs from other sources.
51 Now whilst higher plants generally lack the metabolic pathway for producing fatty acids longer than 18 carbons in length, since the late 1990s research has intensified in the field of transgenic oilseed plants. Transgenic oilseed plants are plants that are sources of short-chain fatty acids which are modified using genetic engineering techniques to introduce foreign genes to enable the production of LC-PUFAs like EPA and DHA. Genes encoding enzymes that enable LC-PUFA production can be sourced from fungi, microalgae and lower plants which are known to produce LC-PUFAs.
52 Let me delve further into the detail and now say something about ω3 LC-PUFA synthesis in plants.
53 All higher plants have the enzymatic complex to synthesize the PUFAs that contain a carbon chain that is 18 carbons long, for example, LA and ALA, but do not possess the enzymes to convert these PUFAs into LC-PUFAs, that is, containing a chain of at least 20 carbon atoms long. The synthesis of LC-PUFAs in higher plants therefore requires the introduction of genes that encode all of the biosynthetic enzymes required to convert either LA into ω6 LC-PUFAs such as AA, or to convert ALA into ω3 LC-PUFAs such as EPA and DHA. These genes can be obtained from a range of organisms that synthesise LC-PUFAs, such as marine bacteria, fungi, protists and microalgae. LC-PUFA synthesis in these organisms involves either aerobic or anaerobic pathways.
54 The aerobic pathway consists of a series of distinct desaturation steps, that is, for each such step the insertion of a C=C double bond into the carbon backbone, and elongation steps, that is, for each such step the insertion of two carbons to the terminal carboxyl group of the fatty acid. Such steps will convert either LA into LC-PUFAs such as AA, EPA and DHA or convert ALA into LC-PUFAs such as EPA and DHA. This conversion of LA or ALA into these LC-PUFAs is shown schematically as follows:
Schematic representation of the conversion of LA or ALA into LC-PUFAs
55 Each of the fatty acids listed in the figure is referred to by its carbon chain length (18, 20 or 22) and the number of C=C double bonds (2, 3, 4, 5 or 6). For instance, LA is referred to as a C18:2 PUFA indicating that it has 18 carbon atoms in its carbon chain and 2 C=C double bonds. Further, the application as filed often includes a reference to whether the PUFA is an omega-6 (ω6) or omega-3 (ω3) PUFA and which carbon atoms along the chain include the C=C double bonds. For example, LA is denoted as a ω6-C18:2Δ9,12 fatty acid, where “Δ9,12” indicates that the 2 C=C double bonds are located at the ninth and twelfth carbon atoms, counting the carbon atoms from the terminal carboxyl end rather than the terminal methyl group. In fatty acid biosynthesis, the “delta” symbol (Δ) indicates the position in the fatty acid at which an enzyme creates a double bond.
56 In the above schematic, the desaturation steps are undertaken by enzymes known as desaturases, and the elongation steps are undertaken by enzymes known as elongases.
57 In the above schematic, the first committed step in the ω6 and ω3 pathways is desaturation of LA and ALA, respectively, by a Δ6-desaturase which introduces a C=C double bond between carbons 6 and 7 of LA and ALA to produce GLA or SDA, respectively.
58 The C18 fatty acids, GLA and SDA, are then elongated by two carbons by a Δ6-elongase to produce the LC-PUFA, di-homo-γ-linolenic acid (DGLA) and eicosatetraenoic acid (ETA), respectively.
59 The products of the Δ6-elongation, DGLA and ETA, may then be desaturated by a Δ5-desaturase which introduces a C=C double bond between carbons 5 and 6 of DGLA and ETA to produce AA and EPA, respectively. This pathway, that is, a Δ6-desaturase followed by a Δ6-elongase and then a Δ5-desaturase, is often referred to as the Δ6-desaturase pathway.
60 An alternative aerobic pathway, often referred to as the alternate Δ9-elongase/Δ8-desaturase pathway, starts with elongation by a Δ9-elongase which adds two carbons to LA and ALA, followed by two successive desaturation reactions. The first desaturation reaction is catalysed by a Δ8-desaturase which introduces a double bond between carbons 8 and 9, producing DGLA and ETA, and the second is catalysed by a Δ5-desaturase, producing AA and EPA. The Δ9-elongase/ Δ8-desaturase pathway is also depicted in the above schematic on the far left hand side and far right hand side, producing the intermediates eicosadienoic acid (EDA) and eicosatrienoic acid (ETrA).
61 In both pathways, EPA is subsequently elongated by two carbons by a Δ5-elongase to DPA. And then DPA may be desaturated by a Δ4-desaturase which introduces a double bond between carbons 4 and 5 to yield DHA.
62 As the above schematic indicates, the ω6 PUFAs include LA and fatty acids derived from LA by desaturation and elongation. The ω6 PUFAs include GLA, 18:3Δ6,9,12, EDA, 20:2Δ11,14, DGLA, 20:3Δ8,11,14 and AA, 20:4Δ5,8,11,14.
63 The ω3 fatty acids include ALA and fatty acids derived from ALA by desaturation and elongation. The ω3 PUFAs include SDA, 18:4Δ6,9,12,15, ETrA, 20:3Δ11,14,17, ETA, 20:4Δ8,11,14,17, EPA, 20:5Δ5,8,11,14,17, DPA, 22:5Δ7,10,13,16,19 and DHA, 22:6Δ4,7,10,13,16,19.
64 I should make two other points at this stage.
65 First, in terms of driving fatty acid biosynthesis, the particular structure of the enzyme that is used in the pathway can have an influence over enzyme functionality and efficiency. For example, variation in the amino acid sequence encoding a Δ6-desaturase may influence the enzyme’s ability to bind to one or both of LA and ALA, as well as the efficiency of conversion to GLA and SDA respectively.
66 Second, a Δ6-desaturase in the context of the above schematic will always utilise LA or ALA or both as substrates to introduce a double bond between carbons 6 and 7 from the carboxyl end of the acyl groups of LA and/or ALA. But an enzyme that has Δ6-desaturase catalytic activity may also have other catalytic activity.
(d) A bit more on substrate specificity and catalysis
67 I said that I would return to say something more on the science concerning substrate specificity. Let me do so now.
68 In evidence before me were only incomplete extracts from Lehninger AL’s Biochemistry (2nd edition, 1975, Worth Publishers Inc (N.Y.)) and Lehninger: Principles of Biochemistry edited by Nelson DL and Cox MM (4th edition, 2005, W.H Freeman and Company) concerning enzymes including their structure and catalytic function. The extracts tendered in part dealt with substrate specificity. It was necessary for me to procure with the assistance of the Court’s Victorian librarian, Mr Michael Coats, other parts of Chapter 6 of the 4th edition in order to put into context the extract that had been given to me. For what it is worth, I was also able to procure and confirm that the current 7th edition (2017) did not substantially differ from the 4th edition as to the matters that I am concerned with; in any event it is well after the relevant date applicable to the construction questions. The following may be distilled from the 2nd and 4th editions, with the 7th edition put to one side.
69 First, generally speaking, one of the notable features of enzymes is their specificity of action when compared with synthetic catalysts or inorganic catalysts. And but for such specificity, one would be swamped with unwanted if not deleterious side reactions and collateral products.
70 Second, unlike most other catalysts, enzymes have very significant catalytic power in dilute aqueous solutions at moderate temperatures and in biological pH ranges.
71 Third, enzymes vary in their specificity. Some will have near absolute specificity for a particular substrate. Others can bind to a whole class of molecules being structurally related substrates, but at different rates. Moreover, as the 2nd edition explained, “substrate molecules generally reflect, by the principle of complementarity, the structure of the active site of the enzyme in two distinctive structural features: (1) the substrate must have a susceptible chemical bond that can be attacked by the enzyme; (2) it usually has some other structural feature required for its binding to the enzyme active site, presumably to position the substrate molecule in the proper geometrical relationship so that the susceptible bond can be attacked.”
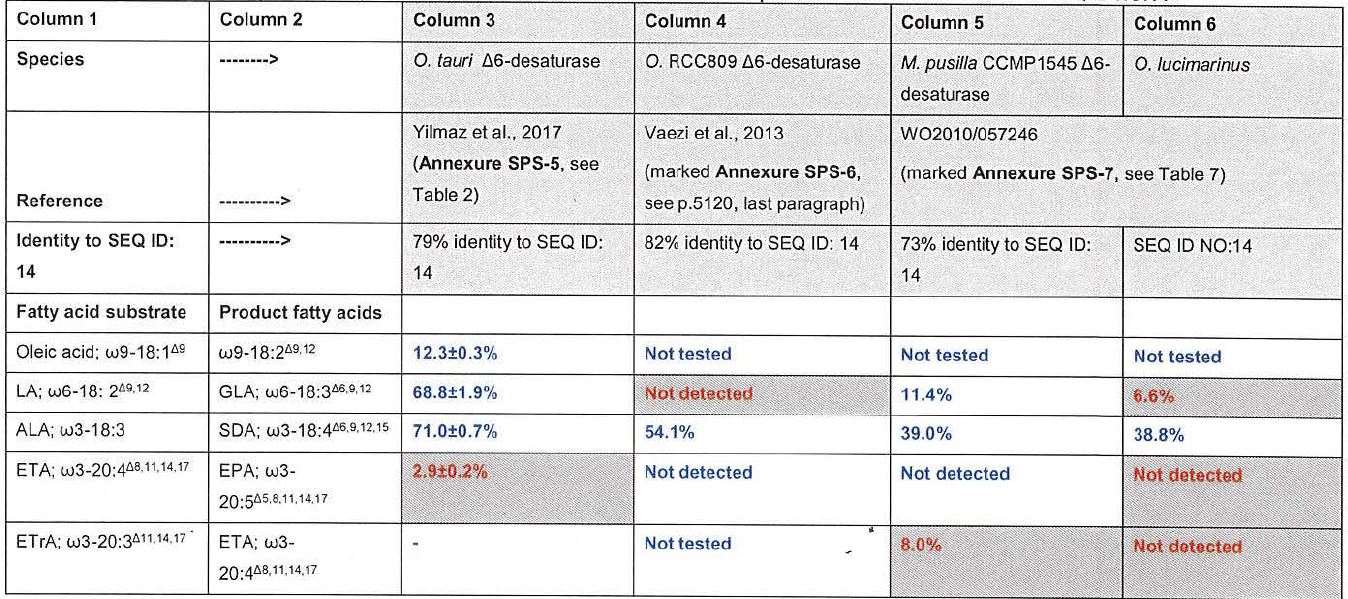
72 Clearly, in the context of the 2nd edition extract that I was given, specificity relates to the question of the enzyme bonding to the substrate. In this context, it is concerned with the ability of the enzyme to discriminate between a substrate and a competing molecule. Let me elaborate further by reference to section 6.2 of the 4th edition and distinguish between the concepts of substrate specificity and catalytic effect, albeit of course related. I have done so because it was not entirely clear to me during the concurrent evidence session that the experts had sufficiently described the relevant concepts to my satisfaction; this of course reveals my own limitations rather than those of the experts. At one stage there appeared to be a blend between specificity and catalytic function and mechanism. Moreover, at one stage it seemed to me that the concepts of substrate and product had also been blended, although I understood that a substrate transformed through intermediate steps to a product by one enzyme could then function as a substrate upon which another enzyme could act, so producing another product, and so on. Further, in terms of substrate specificity as a precursor to catalytic function, the type of bonding/binding was not entirely clear to me. So, it has become necessary to expand on some of these concepts by reference to one of the standard works in the area at the time. I would take the person skilled in the art to have been well aware of the following biochemical concepts when coming to read the specification at the relevant time.
73 In terms of substrates, it is explained:
The distinguishing feature of an enzyme-catalyzed reaction is that it takes place within the confines of a pocket on the enzyme called the active site. The molecule that is bound in the active site and acted upon by the enzyme is called the substrate. The surface of the active site is lined with amino acid residues with substituent groups that bind the substrate and catalyze its chemical transformation. Often, the active site encloses a substrate, sequestering it completely from solution. The enzyme-substrate complex, whose existence was first proposed by Charles-Adolphe Wurtz in 1880, is central to the action of enzymes. It is also the starting point for mathematical treatments that define the kinetic behaviour of enzyme-catalyzed reactions and for theoretical descriptions of enzyme mechanisms (emphasis in original.)
74 The mechanics of catalysis are then explained in the following terms:
A simple enzymatic reaction might be written
E + S ⇌ ES ⇌ EP ⇌ E + P
where E, S, and P represent the enzyme, substrate, and product; ES and EP are transient complexes of the enzyme with the substrate and with the product.
The function of a catalyst is to increase the rate of a reaction. Catalysts do not affect reaction equilibria. Any reaction, such as S ⇌ P, can be described by a reaction coordinate diagram (Fig. 6-2), a picture of the energy changes during the reaction. Energy in biological systems is described in terms of free energy, G. In the coordinate diagram, the free energy of the system is plotted against the progress of the reaction (the reaction coordinate). The starting point for either the forward or the reverse reaction is called the ground state, the contribution to the free energy of the system by an average molecule (S or P) under a given set of conditions. To describe the free-energy changes for reactions, chemists define a standard set of conditions (temperature 298 K; partial pressure of each gas 1 atm, or 101.3 kPa; concentration of each solute 1 M) and express the free-energy change for this reacting system as ΔG, the standard free-energy change. Because biochemical systems commonly involve H+ concentrations far below 1 M, biochemists define a biochemical standard free-energy change, ΔG’, the standard free-energy change at pH 7.0.
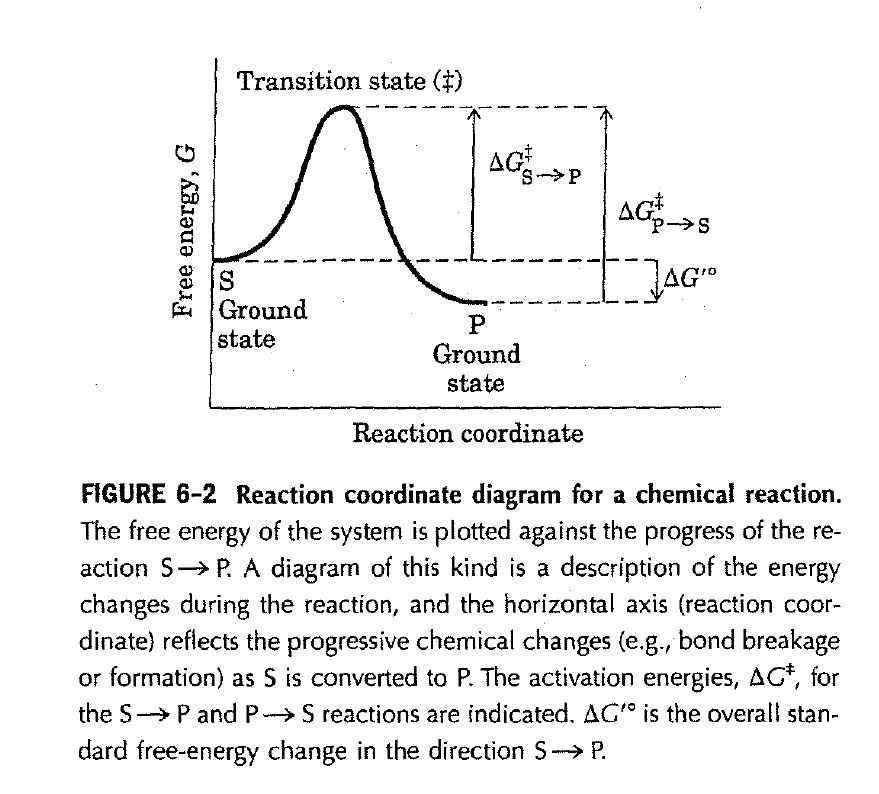
FIGURE 6-2 Reaction coordinate diagram for a chemical reaction.
The free energy of the system is plotted against the progress of the reaction S P. A diagram of this kind is a description of the energy changes during the reaction, and the horizontal axis (reaction coordinate) reflects the progressive chemical changes (e.g., bond breakage or formation) as S is converted to P. The activation energies, ΔG±, for the S P reactions are indicated ΔG’ is the overall standard free-energy change in the direction S P.
The equilibrium between S and P reflects the difference in the free energies of their ground states. In the example shown in Figure 6-2, the free energy of the ground state of P is lower than that of S, so ΔG’ for the reaction is negative and the equilibrium favors P. The position and direction of equilibrium are not affected by any catalyst.
A favorable equilibrium does not mean that the S P conversion will occur at a detectable rate. The rate of a reaction is dependent on an entirely different parameter. There is an energy barrier between S and P: the energy required for alignment of reacting groups, formation of transient unstable charges, bond rearrangements, and other transformations required for the reaction to proceed in either direction. This is illustrated by the energy “hill” in Figure 6-2. To undergo reaction, the molecules must overcome this barrier and therefore must be raised to a higher energy level. At the top of the energy hill is a point at which decay to the S or P state is equally probable (it is downhill either way). This is called the transition state. The transition state is not a chemical species with any significant stability and should not be confused with a reaction intermediate (such as ES or EP). It is simply a fleeting molecular moment in which events such as bond breakage, bond formation, and charge development have proceeded to the precise point at which decay to either substrate or product is equally likely. The difference between the energy levels of the ground state and the transition state is the activation energy, ΔG±. The rate of a reaction reflects this activation energy: a higher activation energy corresponds to a slower reaction. Reaction rates can be increased by raising the temperature, thereby increasing the number of molecules with sufficient energy to overcome the energy barrier. Alternatively, the activation energy can be lowered by adding a catalyst. Catalysts enhance reaction rates by lowering activation energies (emphasis in original).
75 Further, it is explained that any reaction may have several steps, involving the formation and decay of transient chemical species called reaction intermediates. A reaction intermediate is any species on the reaction pathway that has a finite chemical lifetime. When the S ⇌ P reaction is catalyzed by an enzyme, the ES and EP complexes can be considered intermediates, even though S and P are stable chemical species. Further, when several steps occur in a reaction, the overall rate is determined by the step (or steps) with the highest activation energy; this is called the rate-limiting step.
76 The question was then posed as to what was the source of the energy for the dramatic lowering of the activation energies for specific reactions. The answer to the question had two distinct but related parts and was given in the following terms:
The first lies in the rearrangements of covalent bonds during an enzyme-catalyzed reaction. Chemical reactions of many types take place between substrates and enzymes’ functional groups (specific amino acid side chains, metal ions, and coenzymes). Catalytic functional groups on an enzyme may form a transient covalent bond with a substrate and activate it for reaction, or a group may be transiently transferred from the substrate to the enzyme. In many cases, these reactions occur only in the enzyme active site. Covalent interactions between enzymes and substrates lower the activation energy (and thereby accelerate the reaction) by providing an alternative, lower-energy reaction path.
The second part of the explanation lies in the non-covalent interactions between enzyme and substrate. Much of the energy required to lower activation energies is derived from weak, noncovalent interactions between substrate and enzyme. What really sets enzymes apart from most other catalysts is the formation of a specific ES complex. The interaction between substrate and enzyme in this complex is mediated by the same forces that stabilize protein structure, including hydrogen bonds and hydrophobic and ionic interactions. Formation of each weak interaction in the ES complex is accompanied by release of a small amount of free energy that provides a degree of stability to the interaction. The energy derived from enzyme-substrate interaction is called binding energy, ΔGB. Its significance extends beyond a simple stabilization of the enzyme-substrate interaction. Binding energy is a major source of free energy used by enzymes to lower the activation energies of reactions (emphasis in original.)
Two fundamental and interrelated principles provide a general explanation for how enzymes use noncovalent binding energy:
1. Much of the catalytic power of enzymes is ultimately derived from the free energy released in forming many weak bonds and interactions between an enzyme and its substrate. This binding energy contributes to specificity as well as to catalysis.
2. Weak interactions are optimized in the reaction transition state; enzyme active sites are complementary not to the substrates per se but to the transition states through which substrates pass as they are converted to products during an enzymatic reaction.
77 Later it was said:
The same binding energy that provides energy for catalysis also gives an enzyme its specificity, the ability to discriminate between a substrate and a competing molecule. Conceptually, specificity is easy to distinguish from catalysis, but this distinction is much more difficult to make experimentally, because catalysis and specificity arise from the same phenomenon. If an enzyme active site has functional groups arranged optimally to form a variety of weak interactions with a particular substrate in the transition state, the enzyme will not be able to interact to the same degree with any other molecule. For example, if the substrate has a hydroxyl group that forms a hydrogen bond with a specific Glu residue on the enzyme, any molecule lacking a hydroxyl group at that particular position will be a poorer substrate for the enzyme. In addition, any molecule with an extra functional group for which the enzyme has no pocket or binding site is likely to be excluded from the enzyme. In general, specificity is derived from the formation of many weak interactions between the enzyme and its specific substrate molecule. (my emphasis.)
78 So, the mechanism for catalysis is separate from although of course related to the question of specificity. Moreover, as the authors explain, it is the weak interactions between the enzyme and the substrate that provide a substantial driving force for catalysis. And it was explained:
The groups on the substrate that are involved in these weak interactions can be at some distance from the bonds that are broken or changed. The weak interactions formed only in the transition state are those that make the primary contribution to catalysis. The requirement for multiple weak interactions to drive catalysis is one reason why enzymes (and some coenzymes) are so large. An enzyme must provide functional groups for ionic, hydrogen-bond, and other interactions, and also must precisely position these groups so that binding energy is optimized in the transition state. Adequate binding is accomplished most readily by positioning a substrate in a cavity (the active site) where it is effectively removed from water. The size of proteins reflects the need for superstructure to keep interacting groups properly positioned and to keep the cavity from collapsing.
The Evolution Of The Application
79 At this point let me go through the evolution of the application, dealing first with the application as filed, then the application as accepted, and then the proposed amendments to the application as accepted; such amendments in context will then need to be compared with the application as filed.
(a) Application as filed
80 The specification of the application as filed is entitled “Process for the production of polyunsaturated fatty acids in transgenic organisms”.
81 The specification commences (p 1 lines 5 to 13) with the following description of the field of the invention:
The present invention relates to polynucleotides from Ostreococcus lucimarinus which code for desaturases and elongases and which can be employed for the recombinant production of polyunsaturated fatty acids. The invention furthermore relates to vectors, host cells and transgenic nonhuman organisms which comprise the polynucleotides, and to the polypeptides encoded by the polynucleotides. Finally, the invention also relates to production processes for the polyunsaturated fatty acids and for oil, lipid and fatty acid compositions.
82 Ostreococcus lucimarinus (O. lucimarinus) is a species of unicellular algae.
83 Various health benefits of PUFAs (e.g. brain development and function; eye function; cholesterol levels; arthritis) are described (p 1, line 23 to p 3, line 4).
84 It is explained (p 1 lines 15 to 30):
Fatty acids and triacylglycerides have a multiplicity of applications in the food industry, in animal nutrition, in cosmetics and in the pharmacological sector. Depending on whether they are free saturated or unsaturated fatty acids or else triacylglycerides with an elevated content of saturated or unsaturated fatty acids, they are suitable for very different applications. Polyunsaturated fatty acids such as linoleic acid and linolenic acid are essential for mammals, since they cannot be produced by the latter. Polyunsaturated ω3-fatty acids and ω6-fatty acids are therefore an important constituent in animal and human nutrition.
Polyunsaturated long-chain ω3-fatty acids such as eicosapentaenoic acid (= EPA, C20:5Δ5,8,11,14,17) or docosahexaenoic acid (= DHA, C22:6Δ4,7,10,13,16,19) are important components in human nutrition owing to their various roles in health aspects, including the development of the child brain, the functionality of the eyes, the synthesis of hormones and other signal substances, and the prevention of cardiovascular disorders, cancer and diabetes (Poulos, A Lipids 30:1-14, 1995; Horrocks, LA and Yeo YK Pharmacol Res 40:211-225, 1999). This is why there is a demand for the production of polyunsaturated long-chain fatty acids.
85 At page 2, lines 8 to 19 the specification explains that LC-PUFAs, such as EPA, DHA, AA (ARA) and DPA are conventionally obtained from fish and not synthesised in oil producing crops. The following is said:
The various fatty acids and triglycerides are mainly obtained from microorganisms such as Mortierella and Schizochytrium or from oil-producing plants such as soybean, oilseed rape, algae such as Crypthecodinium or Phaeodactylum and others, where they are obtained, as a rule, in the form of their triacylglycerides (= triglycerides = triglycerols). However, they can also be obtained from animals, such as, for example, fish. The free fatty acids are advantageously prepared by hydrolysis. Very long-chain polyunsaturated fatty acids such as DHA, EPA, arachidonic acid (=ARA, C20:4Δ5,8,11,14), dihomo-γ-linolenic acid (C20:3Δ8,11,14) or docosapentaenoic acid (DPA, C22:5Δ7,10,13,16,19) are not synthesized in oil crops such as oilseed rape, soybean, sunflower or safflower. Conventional natural sources of these fatty acids are fish such as herring, salmon, sardine, redfish, eel, carp, trout, halibut, mackerel, zander or tuna, or algae.
86 The specification acknowledges that, “there has been no lack of attempts in the past to make available genes which are involved in the synthesis of fatty acids or triglycerides for the production of oils in various organisms …” (p 3 lines 5 to 7). Such prior art is then described (p 3, line 8 to p 4, line 26).
87 It is then explained (p 4 line 27 to p 5 line 12):
Depending on their desaturation pattern, the polyunsaturated fatty acids can be divided into two large classes, viz. ω6- or ω3-fatty acids, which differ with regard to their metabolic and functional activities.
The starting material for the ω6-metabolic pathway is the fatty acid linoleic acid (18:2Δ9,12) while the ω3-pathway proceeds via linolenic acid (18:3Δ9,12,15). Linolenic acid is formed by the activity of an ω3-desaturase (Tocher et al. 1998, Prog. Lipid Res. 37, 73-117; Domergue et al. 2002, Eur. J. Biochem. 269, 4105-4113).
Mammals, and thus also humans, have no corresponding desaturase activity (Δ12- and ω3-desaturase) and must take up these fatty acids (essential fatty acids) via the food. Starting with these precursors, the physiologically important polyunsaturated fatty acids arachidonic acid (= ARA, 20:4Δ5,8,11,14), an ω6-fatty acid and the two ω3-fatty acids eicosapentaenoic acid (= EPA, 20:5Δ5,8,11,14,17) and docosahexaenoic acid (DHA, 22:6Δ4·7,10,13,17,19) are synthesized via the sequence of desaturase and elongase reactions. The application of ω3-fatty acids shows the therapeutic activity described above in the treatment of cardiovascular diseases (Shimikawa 2001, World Rev. Nutr. Diet. 88, 100-108), inflammations (Calder 2002, Proc. Nutr. Soc. 61, 345-358) and arthritis (Cleland and James 2000, J. Rheumatol. 27, 2305-2307).
88 The specification identifies that it would be advantageous to introduce genes that encode for the enzymes for LC-PUFA biosynthesis in higher plants (p 6 lines 9 to 16):
To this end, it is advantageous to introduce, into oil crops, genes which encode enzymes of the LCPUFA biosynthesis via recombinant methods and to express them therein. These genes encode for example Δ6-desaturases, Δ6-elongases, Δ5-desaturases or Δ4-desaturases. These genes can advantageously be isolated from microorganisms and lower plants which produce LCPUFAs and incorporate them in the membranes or triacylglycerides. Thus, it has already been possible to isolate Δ6-desaturase genes from the moss Physcomitrella patens and Δ6-elongase genes from P. patens and from the nematode C. elegans.
89 The specification acknowledges that such plants, comprising and expressing genes encoding the enzymes necessary for LC-PUFA biosynthesis, have been described in the prior art (p 6 lines 18 to 22), but that they produce LC-PUFAs in “amounts which require further optimization for processing the oils which are present in the plants”.
90 The object of the invention is then stated (p 6, lines 24 to 30):
To make possible the fortification of food and of feed with these polyunsaturated fatty acids, there is therefore a great need for means and measures for a simple inexpensive production of these polyunsaturated fatty acids, specifically in eukaryotic systems. The object of the present invention would therefore be the provision of such means and measures. This object is achieved by the use forms which are described in the patent claims and hereinbelow.
91 The invention the subject of the application is then discussed (p 6 line 31 to p 7 line 2):
The invention, the subject of the present application, is directed to the following:
• a CoA-dependent delta-6-desaturase having the substrate specificity of the delta-6-desaturase shown in SEQ ID NO:14, and
• the above CoA-dependent delta-6-desaturase which has a preference for conversion of alpha linolenic acid compared to linoleic acid.
92 As this passage spans pp 6 and 7 of the application as filed, it has been referred to by the experts as the bridging paragraph. I will also use that description.
93 “CoA-dependent” describes a Δ6-desaturase that predominantly desaturates acyl-Coenzyme A (CoA) bound fatty acid substrates in the cytosolic acyl-CoA pool. SEQ ID NO: 13 is the polynucleotide which encodes the Δ6-desaturase from O. lucimarinus. SEQ ID NO: 14 is the polypeptide sequence encoded by SEQ ID NO: 13, that is, the CoA-dependent Δ6-desaturase polypeptide from O. lucimarinus. Of course, similar polynucleotides may either code for the same polypeptide or for a polypeptide with a similar sequence that preserves the identified biological function of interest. But of course, they may not.
94 As I have said, a substrate is the starting molecule upon which an enzyme acts. An enzyme may bind to and convert multiple substrates. Substrate specificity means the specific substrates, or the number of molecules, that an enzyme binds to at the catalytic site. There is a dispute as to the meaning of “the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14”.
95 The experts have agreed that the words “the above” mean that the statement in the second bullet point of the bridging paragraph must be read together with the statement in the first bullet point of the bridging paragraph. Further, the feature in the first bullet point does not need to include the ALA preference feature disclosed in the second bullet point.
96 The conversion preference referred to means that the CoA-dependent Δ6 desaturase has greater desaturase activity on ALA relative to LA, that is it preferentially converts ALA to SDA over converting LA to GLA.
97 The experts have agreed that the bridging paragraph is the only place in which a conversion preference for ALA or, indeed, any conversion preference is disclosed in the body of the specification of the application as filed. Moreover, they have agreed that it is only disclosed in the limited context of, and as a feature of, a CoA-dependent Δ6-desaturase having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14.
98 The invention described in the bridging paragraph is specifically claimed in claims 1 and 2 respectively of the application as filed.
99 Page 7, lines 4 to 18 of the application as filed then describe a number of matters “[a]ccording to the invention…”. This provides:
According to the invention, the term “polynucleotide” relates to polynucleotides which comprise nucleic acid sequences which code for polypeptides with desaturase or elongase activity. The desaturase or elongase activities are preferably required for the biosynthesis of lipids or fatty acids. Especially preferably, they take the form of the following desaturase or elongase activities: Δ4-desaturase, Δ5-desaturase, Δ5-elongase, Δ6-desaturase, Δ6-elongase or Δ12-desaturase. The desaturases and/or elongases are preferably involved in the synthesis of polyunsaturated fatty acids (PUFAs) and especially preferably in the synthesis of long-chain PUFAs (LCPUFAs). Suitable detection systems for these desaturase or elongase activities are described in the examples or in WO2005/083053. Especially preferably, the above-mentioned activities are, as regards substrate specificities and conversion rates, those of the respective enzymes from Ostreococcus lucimarinus. The specific polynucleotides according to the invention, i.e. the polynucleotides with a nucleic acid sequence as shown in SEQ ID NO: 1, 3, 5, 7, 9, 11, 13 or 15, were obtained from Ostreococcus lucimarinus.
100 Various desaturases and elongases, including Δ4-desaturase activity, Δ5-desaturase activity, Δ5-elongase activity, Δ6-desaturase activity, Δ6-elongase activity or Δ12-desaturase activity, are described. As indicated, it concludes:
Especially preferably, the above-mentioned activities are, as regards substrate specificities and conversion rates, those of the respective enzymes from Ostreococcus lucimarinus.
101 But in dealing with Δ6 desaturases, relevantly this is to be read in the context of the bridging paragraph.
102 The specification then states on p 7 line 20 and p 8 lines 15 to 20:
Therefore, polynucleotides according to the invention are in particular:
…
Polynucleotides which code for a polypeptide with Δ6-desasturase activity and which (i) comprise a nucleic acid sequence as shown in SEQ ID NO:13, (ii) comprise a nucleic acid sequence which codes for a polypeptide as shown in SEQ ID NO:14, (iii) comprise a nucleic acid sequence with at least 72% identity to one of the nucleic acid sequences of (i) or (ii), or (iv) a nucleic acid sequence of a fragment of a nucleic acid from (i), (ii) or (iii).
103 For convenience, and as the experts have done, I will refer to this as the page 8 paragraph.
104 The page 8 paragraph broadly describes polynucleotides which encode a polypeptide having Δ6-desaturase activity. According to the sequence listing, SEQ ID NO: 13 is a polynucleotide which encodes the Δ6-desaturase from O. lucimarinus. From a visual comparison, it would seem that SEQ ID NO: 14 is the polypeptide sequence encoded by SEQ ID NO: 13.
105 Sub-paragraph (i) of the page 8 paragraph provides for polynucleotides that incorporate SEQ ID NO: 13 and which code for a polypeptide having Δ6-desaturase activity.
106 Sub-paragraph (ii) provides for polynucleotides which code for a polypeptide having the amino acid sequence of SEQ ID NO: 14, which is a sequence that has Δ6-desaturase activity. But there would be a pool of polynucleotide sequences that could encode for a protein that has the activity of the protein of SEQ ID NO: 14, given the redundancy in the genetic code.
107 Sub-paragraph (iii) provides for polynucleotides which code for a polypeptide having Δ6-desaturase activity and which have at least 72% identity to those sequences of sub-paragraphs (i) or (ii). Percentage identity would be determined by performing a polynucleotide sequence alignment. On its face this provides for a broader range of polynucleotides than in (i) and (ii).
108 Finally, sub-paragraph (iv) provides for polynucleotides which code for a polypeptide having Δ6-desaturase activity, but which comprise only a fragment of the sequences of (i), (ii) or (iii). This is also on its face a broader range of polynucleotides than in (i) and (ii).
109 In summary, sub-paragraphs (iii) and (iv) of the page 8 paragraph describe a broader range of polynucleotides than sub-paragraphs (i) and (ii) and the polypeptide that is coded for may have a range of differing properties. Now although the experts agree that, taken in isolation, the polynucleotide described at (iii) may not have the substrate specificity of the CoA-dependent Δ6-desaturase shown in SEQ ID NO: 14 or the relevant conversion preference as described in the bridging paragraph, in my view the descriptions at page 7, line 13 and in the page 8 paragraph should be read in the context of the invention disclosed in the bridging paragraph. That is, read as a whole, the polynucleotides described in the page 8 paragraph are examples of polynucleotides coding for the invention described in the bridging paragraph.
110 Further, the invention is also said to relate to a process for the production of a substance which has the formula shown on pages 38 to 39. The formula covers an enormous array of chemical compounds and LC-PUFAs. This aspect of the invention is claimed in dependent claim 9 of the application as filed.
111 Figures 1 to 5 of the specification compare, respectively, the sequence alignment of Δ5- and Δ6-elongase, Δ4-desaturase, Δ5-desaturase, Δ6-desaturase and Δ12-desaturase amino acid sequences from O. lucimarinus and other unicellular algae (p 64 lines 5 to 20). Figure 4 relates to Δ6-desaturase.
112 Figures 6 to 10 show gas-chromatographic determination of fatty acids from yeasts which have been transformed with various plasmids (p 64 line 21 to p 65 line 5).
113 The specification concludes with five examples of “the present invention” (p 65 lines 7 and 8). Examples 1 to 3 relate to general methods of cloning, sequence analysis and lipid extraction. Example 4 relates to cloning and characterisation of elongase genes from O. lucimarinus. Example 5 relates to cloning and characterisation of desaturase genes from O. lucimarinus. Δ6-desaturase with SEQ ID No: 14 (this did refer to SEQ ID No: 30 in the application as filed but “30” has now been corrected to “14”) is characterised and its sequence similarity is compared to Δ6 desaturases from other algae (p 72 line 15 to p 74 line 26). But the activity and substrate specificity is only provided for two Δ5-desaturases and a Δ12-desaturase and not the Δ6 desaturase (pp 74 to 77).
114 The application as filed ends in 15 claims. Claim 1 is the only independent claim. Claims 2 to 8 are product claims that depend on claim 1. Claims 9 to 11 are process claims that ultimately depend on claim 1. Claims 12 to 14 are use claims that ultimately depend on claim 1. Claim 15 is a product by process claim, dependent on claims 9 to 11.
115 Claim 1 of the application as filed is for:
A CoA-dependent delta-6 desaturase having the substrate specificity of the delta-6 desaturase shown in SEQ ID NO: 14.
116 For convenience, and as the CSIRO has submitted, the “substrate specificity of the delta-6 desaturase shown in SEQ ID NO: 14” can be referred to as feature A. It is stipulated in the first bullet point of the bridging paragraph.
117 Dependent claim 2 of the application as filed is for:
The CoA-dependent delta-6 desaturase according to claim 1, wherein the desaturase has a preference for conversion of alpha linolenic acid compared to linoleic acid.
118 Again for convenience, a “preference for conversion of alpha linolenic acid compared to linoleic acid” can be referred to as feature B. It is stipulated in the second bullet point of the bridging paragraph. Accordingly, dependent claim 2 of the application as filed is for a CoA-dependent Δ6-desaturase having both feature A and feature B.
119 Dependent claim 3 of the application as filed is for:
An isolated polynucleotide comprising a nucleic acid sequence coding for the CoA-dependent Delta-6 desaturase according to claim 1.
120 Dependent claim 5 of the application as filed is relevantly for:
A host cell comprising the polynucleotide according to claim 3…
121 Dependent claim 9 of the application as filed is for:
A process for the production of a substance which has the structure shown in the general formula I hereinbelow:
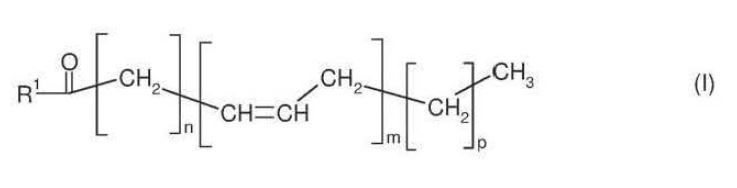
where the variables and substituents are as follows:
R1 = hydroxyl, coenzyme A (thioester), lysophosphatidylcholine, lysophosphatidylethanolamine, lysophosphatidylglycerol, lysodiphosphatidylglycerol, lysophosphatidylserine, lysophosphatidylinositol, sphingo base or a radical of the formula II
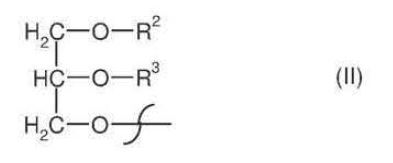
R2 = hydrogen, lysophosphatidylcholine, lysophosphatidylethanolamine, lysophosphatidylglycerol, lysodiphosphatidylglycerol, lysophosphatidylserine, lysophosphatidylinositol or saturated or unsaturated C2-C24-alkylcarbonyl,
R3 = hydrogen, saturated or unsaturated C2-C24-alkylcarbonyl, or R2 and R3 independently of one another are a radical of the formula Ia:
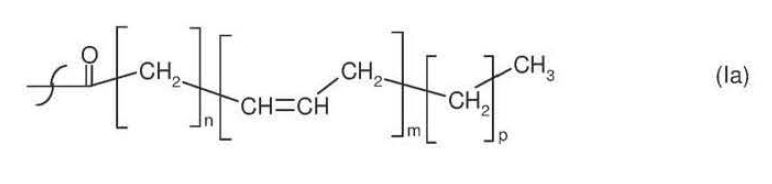
n = 2, 3, 4, 5, 6, 7 or 9, m = 2, 3, 4, 5 or 6 and p = 0 or 3;
and
wherein the process comprises the cultivation of (i) a host cell according to claim 5 or (ii) of a transgenic, nonhuman organism according to claim 7 or claim 8, under conditions which permit the biosynthesis of the substance.
122 A transgenic, nonhuman organism according to claim 7 or claim 8 is one which, inter-alia, comprises an isolated polynucleotide according to claim 3.
123 Dependent claim 12 of the application as filed is for the use of the polynucleotide, or a vector, host cell, transgenic, nonhuman organism comprising the polynucleotide, according to claim 3, for the production of an oil, lipid or fatty acid composition.
124 Finally, it is to be noted that no claim of the application as filed is dependent on claim 2. Further, the application as filed does not claim a process or use that relates to a CoA-dependent Δ6-desaturase having both features A and B. Claim 2 claims a product, being a CoA-dependent Δ6-desaturase having both features.
(b) The application as accepted
125 As I said earlier, the application as filed was amended prior to acceptance on three occasions. Let me note some aspects of the application as accepted incorporating such amendments.
126 The background to the invention described in the application as accepted is identical to that described in the application as filed. But importantly, the bridging paragraph at page 6 line 31 to page 7 line 2 of the application as filed, which defined the CoA-dependent ∆6-desaturase of the invention as having the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14 and a preference for conversion of ALA compared to LA, is not present in the application as accepted.
127 On page 6 line 31 to page 7b line 22 of the application as accepted, the bridging paragraph has been replaced with a description that defines the invention as being directed to a process for the production of substances or compounds of general formula I as follows:
• A process for the production of a substance which has the structure shown in the general formula I hereinbelow:
where the variables and substituents for R1, R2 and R3 are as follows:
…
and
wherein the process comprises the cultivation of:
(i) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence; or
(ii) a transgenic, nonhuman organism comprising:
a) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
b) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence, or
c) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14. wherein the polynucleotide is operatively linked to an expression control sequence,
under conditions which permit the biosynthesis of the substance, and
• use of:
a) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
b) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence,
c) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence, or
d) a transgenic, nonhuman organism comprising:
(i) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
(ii) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence, or
(iii) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14. wherein the polynucleotide is operatively linked to an expression control sequence,
for the production of an oil, lipid or fatty acid composition.
128 The application as accepted defines the invention as being directed to a process for the production of substances or compounds of general formula I, wherein the process requires cultivation of a host cell or a transgenic non-human organism comprising a “polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14”. So, the invention is described as a process for production of a substance by reference to a general formula covering a large number of chemical compounds including LC-PUFAs comprising, and a use of, an isolated polynucleotide having a nucleic acid sequence coding for a CoA-dependent ∆6-desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14.
129 For convenience, the feature “at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14” can be referred to as feature C.
130 Now on its face, in my view a CoA-dependent ∆6-desaturase having feature C is a broader range of polypeptides than a CoA-dependent ∆6-desaturase having feature A, that is, the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14. It describes CoA-dependent ∆6-desaturases that may have a range of different properties and may encompass a large number of variants which do not have feature A.
131 As CSIRO correctly points out, in light of the amendments to the bridging paragraph and the deletion of the corresponding claims, the application as accepted does not refer to feature A, feature B or feature A combined with feature B.
132 Further, the page 8 paragraph remains in the application as accepted and has not been modified from the application as filed.
133 Now as with the application as filed, the application as accepted describes that it relates to polynucleotides from O. lucimarinus which code for desaturases and elongases, and which can be employed for the recombinant production of polyunsaturated fatty acids. Further, the application as accepted describes that the object of the invention is to produce LC-PUFA in eukaryotic systems simply and inexpensively. But unlike the application as filed, the application as accepted claims to have achieved this object by provision of a process for the production of substances or compounds of general formula I. General formula I covers a vast array of chemical compounds and LC-PUFAs, including SDA, EPA, DPA, and DHA, either as free fatty acids or esterified onto a glycerol backbone.
134 The application as accepted has two independent claims that each refer to an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent ∆6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14.
135 Independent claim 1 of the application as accepted claims the process for production of compounds of general formula I, comprising the cultivation of a host cell or a transgenic non-human organism comprising a “polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent ∆6-desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14” as described in the p 6 to p 7b extract set out earlier.
136 Independent claim 5 of the application as accepted claims the use of the isolated polynucleotide, a vector or host cell comprising the isolated polynucleotide, or a transgenic non-human organism comprising the isolated polynucleotide, vector, or host cell for the production of an oil, lipid or fatty acid composition as described in the p 6 to p 7b extract set out earlier.
137 The new description in the specification of the application as accepted (as described in the p 6 to p 7b extract set out earlier) corresponds to accepted process claim 1 and use claim 5.
138 Now claims 1 and 5 and the new description of the application as accepted define the CoA-dependent ∆6-desaturase the subject of the invention as having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 (the O. lucimarinus ∆6-desaturase).
139 But this definition of the claimed CoA-dependent ∆6-desaturase does not equate with, and is not the same as, the requirement of the application as filed for a CoA-dependent ∆6-desaturase having the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14.
140 The reason for this is that altering the nucleotide sequence may change the resulting polypeptide sequence, and could impact on the function of the resulting enzyme, therefore altering the substrate specificity of the enzyme. When nucleotide sequences are altered at multiple positions, the likelihood of the alteration impacting on the enzyme’s function increases. This is because the alteration may impact on the enzyme’s ability to recognise, bind and catalytically act upon particular substrates, therefore altering the enzyme’s substrate specificity.
141 Now I agree with CSIRO that although the application as filed provided a generic description of variants to the polynucleotide sequence encoding SEQ ID NO: 14, this was in the context of the sequence having one or more base substitutions, deletions and or insertions (page 8 line 35 to page 9 line 5); no specific polynucleotide variants were described. Also, it was stated that it was intended that the variants still encode for a polypeptide with the abovementioned biological activity of the respective starting sequence (page 9 lines 3 to 5). So, the invention could include variants, namely, polynucleotide sequences with at least 75% identity, provided that the variants had the same substrate specificity as the starting sequence.
142 And in this regard, the application as filed did not describe which nucleotides could be changed whilst still ensuring that the resulting nucleotide sequence having at least 75% identity to SEQ ID NO: 14 also coded for a polypeptide sequence that had the same substrate specificity of SEQ ID NO: 14.
143 Dr Singh gave evidence that he considered that a CoA-dependent ∆6-desaturase having at least 75% identity to a nucleotide sequence which coded for a polypeptide as shown in SEQ ID NO: 14 was broader than a CoA-dependent ∆6-desaturase having the same substrate specificity to SEQ ID NO: 14 because it included CoA-dependent ∆6-desaturases that did not have the same substrate specificity as SEQ ID NO: 14. I accept that evidence.
144 Indeed, to better illustrate the point that a CoA-dependent ∆6-desaturase having at least 75% identity to a nucleotide sequence which coded for a polypeptide as shown in SEQ ID NO: 14 does not equate with a CoA-dependent ∆6-desaturase having the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14, Dr Singh set out in a Table, which I have reproduced below, a comparison between the known fatty acid substrate specificities of three CoA-dependent ∆6-desaturases – Ostreococcus tauri (O. tauri), O. RCC809 and Micromonas pusilla (M. pusilla) CCMP1545 – and the O. lucimarinus CoA-dependent ∆6-desaturase shown in SEQ ID NO: 14.
Substrate specificity of CoA-dependent ∆6-desaturase variants of the CoA-dependent ∆6-desaturase shown in SEQ ID NO: 14
145 Before discussing the Table, let me note two lesser points. First, each percentage identity was calculated over an aligned region of a specified number of nucleotides. To so calculate, Dr Singh used the NCBI’s BLAST (Basic Local Alignment Search Tool) alignment program to align the nucleotide sequence of each of the O. tauri, O. RCC809 and M. pusilla ∆6-desaturases with the protein coding region of the nucleotide sequence encoding the polypeptide shown in SEQ ID NO: 14, applying the default settings of the standard package. Second, there is one error in the Table. In column 2, the reference to ω9-18:2Δ9,12 should read ∆6, ∆9, omitting ∆12.
146 The results included in the Table were taken from three publications. The first publication was Yilmaz et al., “Determination of Substrate Preferences for Desaturases and Elongases for Production of Docosahexaenoic Acid from Oleic Acid in Engineered Canola”, Lipids (2017) 52 pp 207-222 (Yilmaz). The second publication was International patent application no. WO2010/057246 titled “Enzymes and Method for producing Omega-3 Fatty Acids” (246 Patent). And the third publication was Vaezi et al., “Identification and Functional Characterization of Genes Encoding Omega-3 Polyunsaturated Fatty Acid Biosynthetic Activities from Unicellular Microalgae”, Marine Drugs (2013) 11 pp 5116-5129 (Vaezi).
147 Column 1 of the Table identifies the fatty acid substrates the enzymes act on or bind to, to convert that fatty acid substrate to the fatty acid identified in column 2. This conversion was undertaken experimentally in yeast cells transformed with genetic constructs expressing the desaturase enzymes and which were fed the various fatty acid substrates. The conversion for each fatty acid substrate for each of the three ∆6-desaturase enzymes, compared to the O. lucimarinus ∆6-desaturase, that is, the polypeptide shown in SEQ ID NO: 14, is shown as a percentage of total fatty acids in columns 3 to 6.
148 Yilmaz demonstrated that the O. tauri ∆6-desaturase, which Dr Singh calculated has 79% identity over an aligned region of 1262 nucleotides to nucleotide sequence SEQ ID NO: 13 encoding the polypeptide sequence shown in SEQ ID NO: 14, has a substrate specificity for ETA, as it converts ETA to EPA (a ∆5-desaturase activity) (see Column 3 of the Table above and Table 2 of Yilmaz). In contrast, the O. lucimarinus ∆6-desaturase, that is, the polypeptide shown in SEQ ID NO: 14, had no substrate specificity for ETA, and did not convert ETA to EPA, as EPA was recorded in column 6 as “not detected” (see Table 7 of the 246 Patent). This result according to Dr Singh confirmed that whilst the O. tauri ∆6-desaturase has “at least 75% identity” to the O. lucimarinus ∆6-desaturase of SEQ ID NO: 14, these two enzymes have a different substrate specificity. The nucleotide sequence for the O. tauri ∆6-desaturase gene and its characterisation were published before the priority date of the application as filed by Domergue et al “In vivo characterization of the first acyl-CoA ∆6 desaturase from a member of the plant kingdom, the microalga Ostreococcus tauri” Biochem J (2005) 389:483-449 (Domergue), and its associated Gen Bank sequence listing (Gen Bank Accession No. AY646357.1).
149 Vaezi demonstrated that the O. RCC809 ∆6-desaturase, which Dr Singh calculated has 82% identity over an aligned region of 387-1833 nucleotides to nucleotide sequence SEQ ID NO: 13 encoding the polypeptide sequence shown in SEQ ID NO: 14, does not convert LA to GLA. In contrast, the O. lucimarinus ∆6-desaturase acts on LA.
150 The 246 Patent identified that the M. pusilla CCMP1545 ∆6-desaturase, which Dr Singh calculated has a 73% identity over an aligned region of 1386 nucleotides to nucleotide sequence SEQ ID NO: 13 encoding the polypeptide sequence shown in SEQ ID NO:14, has a substrate specificity for ETrA as it coverts ETrA to ETA (a ∆8-desaturase activity); see Column 5 of the Table. In contrast, the O. lucimarinus ∆6-desaturase was shown not to act on ETrA and therefore had no substrate specificity for ETrA (see Table 7 of the 246 Patent). Although the identity is less than the “at least 75% identity” claimed in the application as accepted, page 8 lines 18 to 19 of the application as filed and of the application as accepted both refer to a nucleic acid sequence with at least 72% identity to SEQ ID NO: 14. According to Dr Singh, this result confirms that whilst the M. pusilla CCMP1545 ∆6-desaturase has “at least 72% identity” to the O. lucimarinus ∆6-desaturase of SEQ ID NO: 14, these two enzymes have a different substrate specificity. Further, this result confirms that variants to the polynucleotide sequence can result in an enzyme that has a different substrate specificity to the original enzyme.
151 In summary, these examples in the evidence demonstrate to my satisfaction that a CoA-dependent ∆6-desaturase having “at least 75% identity” to SEQ ID NO: 14 does not equate to the CoA-dependent ∆6-desaturase having the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14.
152 Let me deal with another matter at this point.
153 The Table also confirms that each of the three comparable enzymes tested O. tauri, O. RCC809 and M. pusilla CCMP1545 have a “preference” for converting ALA over LA as each have a higher percentage of total fatty acids for ALA compared to LA (see columns 3 to 6 of the Table). Further, they have “at least 75% identity” to the O. lucimarinus ∆6-desaturase of SEQ ID NO: 14, or at least 72% identity in the case of M. pusilla, but have a different substrate specificity.
154 So, the CoA-dependent ∆6-desaturase can have a preference for conversion of ALA compared to LA but not have the same substrate specificity as SEQ ID NO: 14.
155 I should say at this point that Dr Stalker did not give any specific evidence in my view which cast any significant doubt on Dr Singh’s Table or the conclusions that could be drawn therefrom, other than to speculate on the existence of possibilities not detected. Now I should say at this point that much of Dr Singh’s material is well after the priority date, but his analysis did assist me to draw some relevant inferences that could justifiably be retrofitted to the relevant earlier timeframe.
156 The application as accepted ends in 9 claims. I should note, inter-alia, that old claims 1 to 3 were removed.
157 Claims 1 and 5 are independent claims. Claim 1 is a process claim. Claim 5 is a use claim. There is no reference in the claims to a CoA-dependent ∆6-desaturase having feature A or a CoA-dependent ∆6-desaturase having features A and B.
158 It is appropriate that I set out the new claims 1 and 5:
1. A process for the production of a substance which has the structure shown in the general formula I hereinbelow:
where the variables and substituents are as follows:
R1 = hydroxyl, coenzyme A (thioester), lysophosphatidylcholine, lysophosphatidylethanolamine, lysophosphatidylglycerol, lysodiphosphatidylglycerol, lysophosphatidylserine, lysophosphatidylinositol, sphingo base or a radical of the formula II
R2 = hydrogen, lysophosphatidylcholine, lysophosphatidylethanolamine, lysophosphatidylglycerol, lysodiphosphatidylglycerol, lysophosphatidylserine, lysophosphatidylinositol or saturated or unsaturated C2-C24-alkylcarbonyl,
R3 = hydrogen, saturated or unsaturated C2-C24-alkylcarbonyl, or R2 and R3 independently of one another are a radical of the formula Ia:
n = 2, 3, 4, 5, 6, 7 or 9, m = 2, 3, 4, 5 or 6 and p = 0 or 3;
and
wherein the process comprises the cultivation of:
(i) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence; or
(ii) a transgenic, nonhuman organism comprising:
a) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
b) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence, or
c) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14. wherein the polynucleotide is operatively linked to an expression control sequence,
under conditions which permit the biosynthesis of the substance.
…
5. The use of:
a) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
b) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence,
c) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14, wherein the polynucleotide is operatively linked to an expression control sequence, or
d) a transgenic, nonhuman organism comprising:
(i) an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO:14,
(ii) a vector comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14, wherein the polynucleotide is operatively linked to an expression control sequence, or
(iii) a host cell comprising an isolated polynucleotide comprising a nucleic acid sequence coding for a CoA-dependent delta-6 desaturase having the substrate specificity of the delta-6 desaturase shown in SEQ ID NO:14. wherein the polynucleotide is operatively linked to an expression control sequence,
for the production of an oil, lipid or fatty acid composition.
(c) The application as proposed to be amended
159 The background to the invention described in the application as proposed to be amended is identical to that described in the application as filed and the application as accepted.
160 The invention described in the application as proposed to be amended adds the following words after the consistory clause at page 6, line 31 to page 7b, which I have set out in the preceding section:
According to an embodiment of the abovementioned process and use, the CoA-dependent desaturase preferentially converts alpha-linolenic acid compared to linoleic acid.
161 The invention claimed in the application as proposed to be amended ends in 11 claims.
162 Claims 1 and 6 are independent claims. Claim 1 is identical to claim 1 in the application as accepted. Claim 6 is identical to claim 5 in the application as accepted.
163 New dependent claim 2 of the application as proposed to be amended is:
The process according to claim 1, wherein the CoA-dependent delta-6 desaturase preferentially converts [ALA] compared to [LA].
164 New dependent claim 7 as proposed to be amended is:
The use according to claim 6, wherein the CoA-dependent delta-6 desaturase preferentially converts [ALA] compared to [LA].
165 None of the claims in the application as proposed to be amended were claimed in the application as filed.
166 As CSIRO has correctly submitted, the application (as accepted) as proposed to be amended would disclose and/or claim a process for the production of, inter-alia, LC-PUFAs comprising an isolated polynucleotide having a nucleic acid sequence coding for a CoA-dependent ∆6-desaturase having feature C combined with feature B (page 6, line 31 to page 7b, line 23 and new claim 2).
167 Further, it would disclose and/or claim a use of an isolated polynucleotide having a nucleic acid sequence coding for a CoA-dependent ∆6-desaturase having feature C combined with feature B (page 6, line 31 to page 7b, line 23 and new claim 7).
168 For convenience I have set out in a schedule to these reasons a table which compares the application as filed with the application as accepted but with the proposed new amendments.
Section 102(1) – Legal Principles
169 The Commissioner must not allow an amendment that is not allowable under s 102 of the Act (s 104(5)). Section 102(1) of the Act as it stood before the Raising the Bar Act prohibited the amendment of a complete specification if the amendment would result in the specification claiming matter not in substance disclosed in the specification as filed. Section 102(1) was in the following terms:
An amendment of a complete specification is not allowable if, as a result of the amendment, the specification would claim matter not in substance disclosed in the specification as filed.
170 Similarly, s 31(1) of the Patents Act 1949 (UK) permitted amendments of a complete specification after acceptance by way of disclaimer, correction or explanation in the following negatively expressed terms:
After the acceptance of a complete specification, no amendment thereof shall be effected except by way of disclaimer, correction or explanation, and no amendment thereof shall be allowed, except for the purpose of correcting an obvious mistake, the effect of which would be that the specification as amended would claim or describe matter not in substance disclosed in the specification before the amendment, or that any claim of the specification as amended would not fall wholly within the scope of a claim of the specification before the amendment.
171 Section 102(1) of the Act (as it then was) was given a broad construction in favour of amendment. The requirement of “in substance disclosed” was treated as very similar to the test for internal fair basis, requiring a “real and reasonably clear disclosure” in the specification as filed of the proposed amendment.
172 In Lockwood Security Products Pty Ltd v Doric Products Pty Ltd (2004) 217 CLR 274 it was said with respect to the internal fair basis test (at [99]):
The inquiry is into what the body of the specification read as a whole discloses as the invention.
173 Further, as the Court explained, merely matching a consistory clause with a claim does not affirmatively establish fair basis. Other parts of the specification may demonstrate the invention to be narrower than the consistory clause.
174 Moreover, even under s 102(1) of the Act as it was, it was not sufficient in and of itself that the effect of the amendment was to narrow the scope of the matter that was claimed to a subset of what was disclosed in the application as filed (Meat & Livestock Australia Ltd v Cargill, Inc (No 2) (2019) 139 IPR 47 at [202], citing AstraZeneca AB v Apotex Pty Ltd (2014) 226 FCR 324 at [244]). In AstraZeneca AB v Apotex Pty Ltd (2014) 226 FCR 324, the plurality said (at [244]):
A very general description of an invention in a specification before amendment might not contain a real and reasonably clear disclosure of more specific embodiments of the invention subsequently disclosed and claimed after amendment. …Whether or not there is a real and reasonably clear disclosure in the specification before amendment of what is claimed in such circumstances is the question that arises in this case.
175 Further, in terms of the nature of the task, as was pointed out by Carr and Goldberg JJ in RGC Mineral Sands Pty Ltd v Wimmera Industrial Minerals Pty Ltd (1998) 89 FCR 458, one first needs to precisely identify the amendment, that is, the amendment of the application as accepted, and second to compare this with the application as filed. As they explained it (at 466):
[Section 102(1)] requires one first to identify precisely what is the amendment. In this case that is done by identifying the difference between the specification as accepted (and as it stood at the hearing of the motion at first instance) on the one hand and, on the other hand, as the specification would read if amended in the manner sought. Then one reads the specification as a whole (as so amended in the manner sought) to see whether as a result of the amendments sought (which must mean by reason of the amendment sought) the specification would claim matter not in substance disclosed in the specification as filed. The subsection focuses on the amendment proposed and it must be that amendment which has the result of pushing the claimed matter over the line defined by the expression “matter not in substance disclosed in the specification as filed”. The key point to keep in mind is, as counsel for the respondent contended (in our view correctly), that the words “as a result of the amendment” are not to be confused with the expression “after the amendment”.
176 Further, the relevant disclosure in the application as filed does not need to be as part of the invention. As they explained (at 468):
Accepting for present purposes (but without deciding) that there are analogies between the fair basing test [then contained in s 45A(2) of the Patents Act 1952 (Cth) and applied by the Full Court in Coopers] and the test for whether matter was in substance disclosed in the specification as filed for the purposes of s 102(1) of the Act, we do not think that too much should be made of the references by Fox J (at the top of 391) and Spender J (at 401) to disclosure “as part of the invention” in the sense of being strictly claimed matter. When their Honours refer to “part of the invention claimed in the provisional specification”, we think it is acceptable and appropriate to treat the word “claimed” as being a reference to “described”: see for example the discussion at 388. So read, Coopers is easily reconciled with the House of Lords decision in the AMP case. In AMP the relevant feature was clearly disclosed in the specification as an optional part of the device. The House of Lords, by a majority, held that it did not matter that it was an optional part. It was sufficient that the optional part was in substance disclosed in the original specification. That is all that is required by s 102(1). In our opinion, to require such disclosure to be part of the invention would be to place an unnecessary and restrictive gloss on a section which cases such as Ethyl Corporation’s Patent [1972] RPC 169 and AMP itself show should be given a liberal interpretation: see Lord Denning MR (at 195) in the former case.
177 The Raising the Bar Act amended s 102(1) of the Act by removing the “in substance disclosed” test.
178 Section 102(1) of the Act now relevantly provides:
(1) An amendment of a complete specification is not allowable if, as a result of the amendment, the specification would claim or disclose matter that extends beyond that disclosed in the following documents taken together:
(a) the complete specification as filed;
(b) other prescribed documents (if any).
179 In March 2009, IP Australia released a consultation paper entitled “Getting the Balance Right: Toward a Stronger and More Efficient IP Rights System”. The consultation paper proposed (at 8, [Proposal 3.2]) that s 40 and s 102 of the Act be amended to:
[E]xplicitly indicate that the requirement for full description is met if the description of the claimed invention was sufficient at the filing date to allow the skilled addressee to perform the invention without undue experimentation.
180 In November 2009, IP Australia issued a second consultation paper entitled “Toward a Stronger and More Efficient IP Rights System”. IP Australia proposed (at 7, [Proposal 1.1.2]) the following changes in relation to s 102(1) of the Act:
Repeal s 102 (1) and replace with a new provision under which an amendment to a complete specification is not allowable if, as a result of the amendment:
• the claims would not be supported by the matter disclosed in the specification at its filing date
• the disclosure contained in the amended specification would go beyond the disclosure contained in the specification at its filing date.
181 This paper stated that the change was to prevent the addition of new matter at a later date and would better align Australian standards for amendment with standards elsewhere in the world.
182 In March 2011, IP Australia initiated a third round of public consultation on IP rights reforms, which included a draft bill and draft explanatory memorandum to give effect, inter-alia, to the proposed change to s 102. The wording of s 102(1)(a) as it presently stands in the Act was included in that draft bill.
183 The explanatory memorandum to the Intellectual Property Laws Amendment (Raising the Bar) Bill 2011 at 60 to 61 explained the rationale for this change to s 102(1) of the Act in the following terms (Item 29):
This item amends the Act to require an applicant to meet the disclosure requirements at the time of filing the complete specification. It is intended to avoid the situation where patent rights accrue in the period before the applicant has fully described their invention.
The Federal Court has recently clarified that an invention need only be fully described at the date of grant. This means that it is possible for an applicant to file an inadequate description that can be subsequently remedied by amendment in order to meet the full description requirements before the date of grant.
This situation is problematic for two reasons.
First, it means that a patentee may gain protection for the period before they have adequately met their obligation to provide the public with a complete disclosure of the invention.
Second, it creates uncertainty for the public and competitors in the period between publication of the patent specification and grant. Without full details of the invention, the public and competitors may not be able to determine where they can safely operate without infringing the patent. They also may not be able to experiment on, or improve the invention.
In contrast to Australia, other countries’ patent laws require the disclosure requirements to be met at filing and do not allow the addition of any material that could not be directly derived from the information in the specification at filing. This includes not allowing the addition of new material to overcome an objection of lack of full description. For the reasons above, this result is preferred to the current Australian position.
The item introduces a provision preventing amendment of a complete patent specification after filing to add new matter that would go beyond the disclosure contained in the specification at its filing date. An applicant would not be able to amend the specification to add any material that the hypothetical skilled person could not directly derive by reading the information in the specification as filed.
(Citation omitted.)
184 During the second reading speech of that Bill, the Minister for Innovation, Industry, Science and Research stated (Commonwealth, Parliamentary Debates, Senate, 22 June 2011, 3485 at 3486) that, inter-alia, the amended provisions were intended to mirror other jurisdictions, such as the UK and Europe, and that it was intended that Australian courts would have regard to the developments of case law in those jurisdictions when interpreting the new provisions.
185 In summary, it would seem that the Raising the Bar Act aligns the approach taken under s 102(1) of the Act with the approach taken by the UK and Europe concerning the dimension of amendment that I am discussing.
186 Let me make some other points.
187 First, the language “claim or disclose matter that extends beyond that disclosed” was previously framed as “claim matter not in substance disclosed”. On its face the addition of the words “or disclose” has a broader purview than the invention defined by the claim. Whether it makes a real difference is another question.
188 Second, there is no express explanation in the explanatory memorandum as to the intended effect of deleting the words “in substance” and replacing them with “extends beyond” (at 60 and 61). But what the explanatory memorandum does emphasise is that the “full description” requirement in s 40(2)(a) should not be able to be satisfied by amending the complete specification as filed, and the explanatory memorandum noted that the full description requirement had also been strengthened by the proposed amendments. In relation to the intended operation of the strengthened s 40(2)(a), the explanatory memorandum said of the amendment (at 46):
This is intended to align the disclosure requirement with that applying in other jurisdictions with the effect that sufficient information must be provided to enable the whole width of the claimed invention to be performed by the skilled person without undue burden, or the need for further invention. This more clearly reflects a fundamental principle of the patent system: in exchange for the exclusive rights given to the patentee, the patentee must share with the public the information necessary to make and use the invention.
189 So, this suggests that there may be impermissible added matter if the amendment disclosed new information that was material to the implementation of the invention, that is, material to satisfying the requirements of s 40(2)(a).
190 Moreover, in relation to the amendments to ss 40(2) and 40(3), the explanatory memorandum disclosed an intention that the operation of those provisions be as close as practicable to that given to the corresponding provisions in UK legislation and the European Patent Convention (EPC).
(a) Patents Act 1977 (UK) and EPC
191 In my view, the requirements in the UK provide relevant guidance as to the scope of s 102(1).
192 The present wording of s 102(1) now corresponds more closely with the language of s 76(2) of the Patents Act 1977 (UK), which in turn is based on article 123(2) of the EPC. As I have indicated, the old wording of s 102(1) corresponded more closely with s 31(1) of the 1949 UK Act.
193 Section 76(2) of the 1977 UK Act (in its form since 7 January 1991) provides:
No amendment of an application for a patent shall be allowed … if it results in the application disclosing matter extending beyond that disclosed in the application as filed.
194 Article 123(2) of the EPC provides:
The European patent application or European patent may not be amended in such a way that it contains subject-matter which extends beyond the content of the application as filed.
195 For present purposes it may be accepted that s 76(2) of the 1977 UK Act has the same meaning as article 123(2) of the EPC. Section 76(3)(a) effectively imposes the same requirement for amendments to granted UK patents.
196 The construction to be given to s 76(2) has been described in Terrell on The Law of Patents, 14th ed (at [7.65]) (see also the 18th ed at [15-40]) as follows:
[T]he section is to be interpreted strictly and the crucial question is ‘whether any subject matter relevant to the invention has been added whether by deletion or addition’…
These decisions and the others referred to in the footnotes show that, apart from disclaiming amendments which limit the monopoly claimed and amendments to correct an obvious mistake, the power to amend is extremely limited and it is difficult to envisage how any amendment to make good any inadequacies of description or to introduce intermediate generalisations can be made.
(Citations omitted.)
197 Although any construction question must begin and end with the statutory text, two conceptual themes permeate the UK authorities which it is convenient to describe and elaborate on. The first concept is “added matter”. The second concept is “intermediate generalisation”, although it is a sub-set of “added matter”. Neither of these concepts are expressed in terms in the statutory language.
Added matter
198 The word “matter” is part of the statutory language, although it is not defined. But clearly it is not just limited to inventive concepts. And if one is dealing with a claim, it would include both form and structure as well as inventive concepts.
199 In Southco Inc v Dzus Fastener Europe Ltd [1990] RPC 587, Aldous J observed (at 616):
What the Act is seeking to prevent is a patentee altering his claims in such a way that they claim a different invention from that which it disclosed in the application. Thus, provided the invention in the amended claim is disclosed in the application when read as a whole, it will not offend against section 76.
200 And in that context in dealing with an amendment pre-grant, the central question does not turn on determining whether it has the effect of widening or narrowing the monopoly claimed.
201 In Bonzel v Intervention Ltd (No 3) [1991] RPC 553, Aldous J stated that the test for examining whether an amendment involved added subject matter was threefold, being (at 574):
(1) To ascertain through the eyes of the skilled addressee what is disclosed, both explicitly and implicitly in the application.
(2) To do the same in respect of the patent as granted.
(3) To compare the two disclosures and decide whether any subject matter relevant to the invention has been added whether by deletion or addition. The comparison is strict in the sense that subject matter will be added unless such matter is clearly and unambiguously disclosed in the application either explicitly or implicitly.
202 This test was approved by Jacob J in Richardson-Vicks Inc’s Patent [1995] RPC 568, where his Honour observed (at 576):
I think the test of added matter is whether a skilled man would, upon looking at the amended specification, learn anything about the invention which he could not learn from the unamended specification.
203 In European Central Bank v Document Security Systems Inc [2007] EWHC 600 (Pat), Kitchen J elaborated on Bonzel (at [97] to [102]) and made a number of points most of which, with respect, I entirely agree. First, the claims form part of the disclosure “though clearly not everything which falls within the scope of the claims is necessarily disclosed”. Second, I must carry out the exercise through the eyes of the skilled addressee who would approach the task with the benefit of the common general knowledge. Third, the two disclosures must be compared to see whether any subject matter relevant to the invention has been added. And this comparison is a strict one. Subject matter will be added “unless it is clearly and unambiguously disclosed in the application as filed.” Moreover, it is appropriate to consider what has been disclosed both explicitly and implicitly. Fourth, what I am considering and applying is not an obviousness test. As Kitchen J described it, a patentee “is not permitted to add matter by amendment which would have been obvious to the skilled person from the application.”
204 The statements in Bonzel, Richardson-Vicks and European Central Bank that I have set out have been approved directly or indirectly in Vector Corporation v Glatt Air Techniques Ltd [2008] RPC 10 at [4] to [8] and Napp Pharmaceutical Holdings Ltd v Ratiopharm GmbH [2009] RPC 18 at [69] to [71].
Intermediate generalisations
205 An amendment is not allowable if it takes a feature which is only disclosed in a particular context and seeks to introduce it into a claim deprived of that context. In the United Kingdom, this is described as an “intermediate generalisation” and is an established aspect of “added matter”.
206 In Palmaz’s European Patents (UK) [1999] RPC 47 Pumfrey J described the concept of “intermediate generalisation” as follows (at 71):
If the specification discloses distinct sub-classes of the overall inventive concept, then it should be possible to amend down to one or other of those sub-classes, whether or not they are presented as inventively distinct in the specification before amendment. The difficulty comes when it is sought to take features which are only disclosed in a particular context and which are not disclosed as having any inventive significance and introduce them into the claim deprived of that context. This is a process sometimes called ‘intermediate generalisation’.
207 Palmaz involved an arterial stent. The specification as filed disclosed second bars of the stent, “extending on the circumference of a circle whose plane is perpendicular to longitudinal axis…” of the stent, but only in the context of the adjacent first bars of the stent being parallel to that axis. The amendment sought to add, as a feature of the independent claim, reference to the second bars of the stent “extending on the circumference of a circle whose plane is perpendicular to longitudinal axis…”, but impermissibly without a requirement of the first bars being parallel to that axis. His Honour disallowed the amendment, observing (at 71):
It seems to me that this amendment represents the selection of a particular feature, whose significance is nowhere disclosed, and its incorporation into the inventive concept shorn of its original context. This feature is for the first time suggested to have technical significance whether or not in combination with bars parallel to the axis of the stent, and in my view this is an addition of matter to the specification.
208 Pumfrey J’s decision was upheld on appeal (Palmaz’s European Patents (UK) [2000] RPC 631) and his conceptual treatment of intermediate generalisations was endorsed in LG Philips LCD Co Ltd v Tatung (UK) Ltd [2007] RPC 21 at [31], [32] and [42] and Vector v Glatt at [9].
209 The decision in Palmaz was explained in Smith & Nephew PLC v Convatec Technologies Inc [2013] RPC 8 by Judge Birss QC in the following terms (at [88]):
Although the words of Pumfrey J in Palmaz and approved in Vector v Glatt refer to features not being disclosed ‘as having any inventive significance’, I do not understand that to mean that only features expressly asserted as being inventive in the specification may be added to a claim by amendment. A patentee does not have to repeatedly assert, with every line of the disclosure, that he thinks each point is or may be inventive. On the other hand of course, the context is crucial and a patentee cannot extract features disclosed in one context and introduce them into a claim stripped of their context.
210 In Nokia Corporation v IPCom GmbH & Co KG (No 3) [2013] RPC 5, Kitchin LJ explained intermediate generalisations (at [56] to [60]) as follows:
Turning to intermediate generalisation, this occurs when a feature is taken from a specific embodiment, stripped of its context and then introduced into the claim in circumstances where it would not be apparent to the skilled person that it has any general applicability to the invention.
Particular care must be taken when a claim is restricted to some but not all of the features of a preferred embodiment, as the TBA explained in decision T 0025/03 at point 3.3 … “if a claim is restricted to a preferred embodiment, it is normally not admissible under Article 123(2) EPC to extract isolated features from a set of features which have originally been disclosed in combination for that embodiment. Such kind of amendment would only be justified in the absence of any clearly recognisable functional or structural relationship among said features…” So also, in decision T 0284/94 the TBA explained … that a careful examination is necessary to establish whether the incorporation into a claim of isolated technical features, having a literal basis of disclosure but in a specific technical context, results in a combination of technical features which is clearly derivable from the application as filed, and the technical function of which contributes to the solution of a recognisable problem. Moreover, it must be clear beyond doubt that the subject matter of the amended claim provides a complete solution to a technical problem unambiguously recognisable from the application.
It follows that it is not permissible to introduce into a claim a feature taken from a specific embodiment unless the skilled person would understand that the other features of the embodiment are not necessary to carry out the claimed invention. Put another way, it must be apparent to the skilled person that the selected feature is generally applicable to the claimed invention absent the other features of that embodiment.
Ultimately the key question is once again whether the amendment presents the skilled person with new information about the invention which is not directly and unambiguously apparent from the original disclosure. If it does then the amendment is not permissible.
211 In summary, an amendment utilising a feature may amount to added subject matter, such that it is not relevantly disclosed, even where there is a literal reference to such a feature in the specification as filed. An example of where this may occur is where by reason of the amendment, a feature that was not suggested to be significant is, for the first time, suggested to have a technical significance. Such a suggestion of technical significance might arise where a feature is taken from a specific embodiment, and introduced into a claim in a manner that is stripped from relevant context provided by that embodiment. To avoid a conclusion of added subject matter, the skilled person must have appreciated from the complete specification as filed that the selected feature was more generally applicable. This may involve asking whether the skilled person is being presented with any new information about the invention which is not directly and unambiguously apparent from the original disclosure. Further, even if a feature was suggested to be technically significant, an impermissible intermediate generalisation may take place if by the amendment the feature is used in a manner significantly different from its original context.
212 Further, what constitutes an impermissible addition of subject matter by way of an “intermediate generalisation” is to be distinguished from an amendment that claims a sub-class of an inventive concept, whether or not it is presented as inventively distinct in the specification as filed. This is consistent also with established practice in relation to patent claiming, where a patent will routinely include cascading claims directed to progressively narrower characterisations of the disclosed invention. A patent applicant is not limited to a single patent claim, directed to a preferred embodiment.
213 Further, there may be an impermissible intermediate generalisation where a new combination of features is sought to be created by the proposed amendment which was not apparent in the application as filed. As has been pointed out (Smith & Nephew PLC at [77]), the question is “whether the features combined in the amended claim were disclosed as a combination in the application [as filed]” (original emphasis).
(b) Australian position
214 I see no good reason not to follow the UK authorities to the extent of applying analogous concepts to the construction of the present form of s 102(1). In summary the following may be noted.
215 First, as has been pointed out, the test is a strict one.
216 Second, subject matter will be impermissibly added unless the matter is clearly and unambiguously disclosed in the application as filed.
217 Third, the required disclosure may be express or implied, but on any view must be clearly and unambiguously so. In this regard, a patent applicant is not permitted to add by amendment matter simply because it would have been obvious to the skilled person.
218 Fourth, context is important. A patent applicant cannot extract features disclosed in one context and introduce them into a specification stripped of that context. So, the concept of intermediate generalisation as discussed in the UK authorities applies to s 102(1) in its construction and application.
219 Indeed on this topic, I would note that even under the prior form of s 102(1) that an “intermediate generalisation” was seen as a form of prohibited amendment. In Les Laboratoires Servier v Apotex Pty Ltd (2010) 273 ALR 630 the specification stated that the invention related to a crystalline form of a salt and a process for its preparation. In describing the process, the specification stated that the salt was obtained in the form of “individual needles”, but there was no mention of individual needles in the earlier description of the product invention. The patentee sought to amend the specification to include claims to the product in the form of “individual needles”. Emmett J held that this feature could not be “linked back” and introduced an additional limitation of the product invention. Emmett J observed (at [25]), Kenny and Stone JJ (at [72]) not deciding,:
The proposed additional claims, incorporating the additional limitation of the product and the process, by reference to individual needles, are not fairly based on the matter contained in the body of the Specification. As a matter of construction of the Specification, the passage that follows the description of the various embodiments of the process invention should not be read back into the description of the product invention. The former does not, as Servier would have it and the primary judge held, link back to the latter. The proposed amendments are not allowable under s 102.
220 Fifth, as to Australian authority, s 102(1) in its present form was recently referred to in Pilkin v Sony Australia Limited (No 2) [2019] FCA 980 in the context of a summary dismissal application and in Cytec Industries Inc v Nalco Company [2019] FCA 1800. Cytec involved an unopposed application and Burley J expressly exercised restraint by declining to analyse in any detail the change effected to s 102(1) by the Raising the Bar Act. And as to Pilkin, it is apparent from the reasoning of Rares J that the significance, if any, of the relevant change was not to the fore.
221 Are the requirements of s 102(1) now stricter? If there is a difference in scope, then the distinction may be subtle. In Palmaz Pumfrey J used the phrase “in substance disclosed”, which was the language used in the 1949 UK Act (at 71), in considering the issue of added matter. Further, relevant editions of Terrell do not refer to any consideration of the impact of the equivalent change in wording from the 1949 UK Act to the 1977 UK Act. Indeed, in the 13th edition of Terrell, published in 1982, the editors thought that the then new s 76(2) would likely be construed “more or less the same as the requirement under the old law that there must be a fair basis for the amendment in the unamended specification” (at [8.64]). And they referred to the issue of “intermediate generalisation” as an issue that presented difficulty under the 1949 UK Act, and would likely continue to be an area of difficulty. But admittedly they also postulated that the new wording might be seen to liberalise the nature of amendments that would be possible.
222 I do not propose to indulge in any disquisition concerning the differences, if any, between the scope of the old s 102(1) and the new s 102(1). As a trial judge that is not my task. And to do so runs the risk of getting lost in some jurisprudential twilight zone outside the dimensions of the documents and the science that I need to consider. Suffice it to say that I am applying s 102(1) as it is, rather than what it was. Moreover, as I have said, UK authorities provide suitable guidance.
BASF’s Arguments
223 BASF says that the relevant amendments including the added new claims 2 and 7 are narrowing amendments.
224 Now in one sense this is true if one compares them with the application as accepted. But of course the question is whether as a result of the amendment, the amended specification would claim or disclose matter that extends beyond that disclosed in the complete specification as filed. So it is not really the issue to compare just the proposed amendments with the application as accepted.
225 BASF’s primary argument, which it says does not require me to resolve what substrate specificity means as used in the bridging paragraph, is that there is disclosure in the page 8 paragraph of a polynucleotide coding for a Δ6 desaturase having the amino acid sequence disclosed as SEQ ID NO: 14, and of certain polynucleotides within specified ranges of sequence homology. Further, it says that the bridging paragraph discloses that the SEQ ID NO: 14 polypeptide preferentially converts ALA compared to LA. I should say now that I do not agree with this last proposition and will return to this later.
226 BASF says that the skilled addressee therefore understands from the complete specification as filed that they have available to them:
(a) Δ6 desaturases that will act in the biosynthetic pathway being addressed by the application; and
(b) as a subset of the above, Δ6 desaturases that will preferentially convert ALA, and will thereby produce more ω3 long-chain fatty acids as compared with ω6 long-chain fatty acids.
227 BASF says that this provides the necessary disclosure for the amendments. Further, it says that the skilled addressee understands that polynucleotides having similar sequence homologies will likely produce polypeptides that preserve those relevant biological functions.
228 Now in support of the primary argument, BASF put four factual propositions.
229 First, the skilled person has available to them a Δ6 desaturase (SEQ ID NO: 14) capable of acting on LA and ALA within the relevant biosynthetic pathway, a schematic for which I have set out earlier in my reasons. It says that the skilled person understands from the context of the complete specification as filed that SEQ ID NO: 14 is being disclosed as a Δ6 desaturase in respect of LA and ALA.
230 Second, it says that the skilled person has available to them a Δ6 desaturase (SEQ ID NO: 14) which is capable of acting on LA and ALA within the schematic for the biosynthetic pathway that I have set out earlier and has the property that the desaturase preferentially converts ALA compared to LA. It says that the skilled person understands from the second bullet point of the bridging paragraph that SEQ ID NO: 14 preferentially converts ALA compared to LA. This proposition is contestable concerning SEQ ID NO: 14, a topic to which I will return.
231 Third, it says that the skilled person understands that variations of the polynucleotide SEQ ID NO: 13 may produce polypeptides that retain the above two properties/functions, namely, Δ6 desaturase activity on LA and ALA, and substrate preference for ALA.
232 Now BASF accepts that any variation of a polynucleotide sequence may potentially result in a polypeptide with changed function. And the more a polynucleotide sequence is changed, the more likely it is that biological functions may also change. However, it says that changes in the polynucleotide sequence may preserve biological function. That includes the two biological functions in question.
233 Fourth, it says that the skilled person understands that variations of the polynucleotide SEQ ID NO: 13 may produce polypeptides that retain the above two properties (Δ6 desaturase activity on LA and ALA, and substrate preference for ALA), whilst varying the substrate specificity as more narrowly construed.
234 It says that the properties of substrate specificity and conversion preference are unrelated, and may vary independently of each other as the coding sequence is varied.
235 BASF’s secondary argument is ancillary to its primary argument. BASF says that the bridging paragraph should not be read as limiting the scope of the disclosure made by the complete specification as filed.
236 BASF submits that the bridging paragraph has a broader construction than that contended for by CSIRO.
237 CSIRO submits that the bridging paragraph discloses what it terms as a CoA dependent Δ6-desaturase having feature A (substrate specificity) combined with feature B (conversion preference for ALA). But BASF says that CSIRO’s features A and B from the bridging paragraph are not combined in the sense of a combination of physical integers. They each describe properties of an individual polypeptide. Further, BASF says that features A and B are not features that, applying CSIRO’s narrow construction of feature A, combine in some way to achieve a relevant specific result.
238 Now the first bullet point of the bridging paragraph says that the invention is directed to a CoA-dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14. But BASF says that this language does not describe the relevant desaturases by reference to either a polypeptide sequence or a polynucleotide sequence.
239 So, it says that whatever the correct meaning is of substrate specificity, the first bullet point sets out a purely functional description. That is, at least in theory, a polypeptide could meet that description whilst having a very different sequence from SEQ ID NO: 14. It could capture subject matter that owed nothing to the development made by the inventors.
240 According to BASF, that is a factor that points away from the bridging paragraph itself strictly delineating the scope of the disclosure made by the complete specification as filed.
241 Further, BASF says that what is relevant about the Δ6-desaturase discovered by the inventors is not that it was able to introduce a desaturated bond at the sixth bond position in a PUFA, but that it was able to do this in respect of two specific substrates being ALA and LA. Those are the two substrates that required a Δ6 desaturation step as part of the biosynthetic pathway to which the application is directed.
242 BASF therefore submits that, read in context, the first bullet point of the bridging paragraph is simply explaining that the invention is not directed to a Δ6 desaturase per se, but is directed to one that acts on ALA and LA, which is what SEQ ID NO: 14 acts on.
243 Now BASF referred to Dr Singh’s narrower view, which is that the substrate specificity looks to every relevant substrate in the biosynthetic pathway under discussion that the enzyme acts on, including where its activity differs. And he gave as an example Δ5-desaturase activity. But BASF says that Dr Singh’s approach makes redundant the two references to Δ6 desaturase in the first bullet point of the bridging paragraph.
244 Moreover, BASF says that there is no other reference in the complete specification as filed to substrate specificity, and no indication that the patent applicant intended to limit the scope of what it was disclosing in the way contended for by CSIRO.
245 BASF says that the complete specification as filed describes the invention broadly, including by reference to various desaturases and elongases, and including by reference to the Δ6 desaturase in the page 8 paragraph.
246 Further, it says that the bridging paragraph does not purport to and is not apt to define an invention as it contains a functional limitation only, and does not set out a scope referable to a polynucleotide sequence or polypeptide sequence. It also says that it is standard patenting practice to claim technical subject matter in this area by reference to sequence homologies.
247 Further, it says that the bridging paragraph does not purport to and is not apt to define an invention, even if “substrate specificity” is narrowly construed.
248 Further, it says that the two bullet points of the bridging paragraph do not define a relevant combination. They do not cooperate, in the sense that they are not both necessary, to achieve some unitary effect or outcome, or solve some identified specific technical problem.
249 Further, it says that a variation in substrate specificity (as narrowly construed) does not necessarily result in reduced yield by reason of unwanted side reactions. The complete specification as filed does not include any discussion of side reactions. In particular, there is no disclosure about side reactions for the SEQ ID NO: 14 enzyme, and no disclosure that it has an optimised level of side reactions. Moreover, it says that if a change in substrate specificity from that of SEQ ID NO: 14 resulted in more side reactions, this might have no impact on yield. There would be no impact if the comparative rate of the side reaction was very low. Further, a change in substrate specificity might result in a side reaction that is then used up in the biosynthetic pathway. Further, a change in substrate specificity might result in fewer side reactions where the enzyme binds to and catalyses fewer substrates than SEQ ID NO: 14.
250 Further, the skilled person would not understand the preferential conversion of ALA to be a desirable property of a Δ6 desaturase only if the substrate specificity of SEQ ID NO: 14 is preserved.
251 BASF’s tertiary argument is that even if the first bullet point of the bridging paragraph is able to narrow the disclosure of the complete specification as filed within the meaning of s 102(1), “substrate specificity” read in context only refers to Δ6 desaturase activity on LA and ALA. So, “substrate specificity”, read in context, refers to the ability of the Δ6-desaturase to act on each of LA and ALA. These are the two substrates that the Δ6-desaturase must act on in the relevant biosynthetic pathway that the application is addressing. BASF says that such an interpretation makes practical sense. It is reinforcing that the Δ6-desaturase must act as a Δ6-desaturase for the relevant substrates. Contrastingly, it says that CSIRO advances no practical explanation as to why the patent applicant would be using the words “substrate specificity” in the way CSIRO contends for.
252 As to the tertiary argument, which is that even if the first bullet point of the bridging paragraph is able to narrow the disclosure, “substrate specificity” only refers to Δ6 desaturase activity on LA and ALA, BASF has put four factual propositions.
253 First, it says that substrate specificity does not refer to the total number of molecules that can bind to an enzyme. It says that there is no definition in either of the 2nd or 4th editions of Lehninger that supports such a definition. Instead the references to specificity are each made by reference to a particular context of candidate substrates. Further, it says that each of the uses of that term in journal articles in evidence was in respect of a limited set of tested substrates only.
254 Second, it says that substrate specificity can have different meanings in different contexts. It says that Dr Singh accepted in his affidavits that substrate specificity required catalytic activity, but that in the joint expert report he had said that substrate specificity could include substrates that bound to the enzyme but were not catalytically converted. Moreover, rather than considering any molecule on which the enzyme might act, Dr Singh inferred at a minimum a limitation that it be referable to fatty acids. Further, it says that Dr Singh considered that, in context, substrate specificity could mean that the Δ6 desaturase meeting the description of the first bullet point could act on more substrates than SEQ ID NO: 14. This suggested a further variation in the meaning of substrate specificity, and could potentially result in more side reactions because more substrates were acted upon.
255 Third, BASF says that a requirement of identical substrate specificity applying Dr Singh’s definition would not provide a workable limitation of the disclosed invention. The Δ6 desaturase is intended to be introduced into a plant, and the complete specification gives many examples of such plants. The plants into which the Δ6 coding sequence is to be introduced may produce a range of different fatty acids. And whether the proposed enzyme would bind to or act on such fatty acids could only be known by testing. But the measured substrate specificity would depend on the limits of detection for the relevant equipment being used. Further, the measured substrate specificity would be dependent on the particular pool of substrates that were tested.
256 Fourth, BASF says that at least one reasonable interpretation of “substrate specificity” in the context of the bridging paragraph is to require the Δ6 desaturase to act on LA and ALA as is the case for SEQ ID NO: 14. That was Dr Stalker’s view.
Analysis
257 Let me first deal with three topics, namely:
(a) substrate specificity;
(b) the preference for conversion of ALA compared to LA; and
(c) the page 8 paragraph.
258 It is convenient to deal with these topics in this order given the flow of the application as filed. I will draw the threads together in a later section. For notation purposes, “delta” has been changed to “Δ” in any direct quote from hereon.
259 For present purposes, I adopt what I said in Meat and Livestock (No 1) at [213] to [220] concerning the applicable principles of construction and the relevant lens of the skilled addressee. I should also say that where and to the extent that it matters, I have preferred Dr Singh’s evidence over Dr Stalker’s evidence on areas of difference, albeit that Dr Singh was not an independent expert. My impression was that he had the slight edge in terms of depth of detail on the precise questions that I was interested in. But of course I accept that the expert evidence can only have a limited role in the present context. And the question as to what was disclosed in the application as filed and the relevant comparison are ultimately matters for me applying the relevant lens.
(a) Substrate specificity
260 In my view, applying the relevant lens and generally speaking, substrate specificity of enzymes addresses the substrates that can bind to an enzyme site such that all of the substrates binding to that site could potentially be converted into a product. Enzymes can have an absolute specificity, that is, bind to one specific molecule and produce one product, or bind to a class of structurally similar molecules and produce a variety of products. But in the context of fatty acid synthesis, being the subject of the application as filed, it can be said that the substrate specificity of an enzyme is the range of fatty acid substrates that the enzyme can bind to and potentially convert to another fatty acid or product(s) and possibly other related molecules.
261 Now the CoA-dependent Δ6-desaturase described in the application as filed is defined functionally as having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14. The Δ6 desaturase sequence shown in SEQ ID NO: 14 is the sequence of the CoA dependent Δ6-desaturase polypeptide from O. lucimarinus. The reference to a CoA-dependent Δ6-desaturase in the application as filed describes a Δ6-desaturase that predominantly desaturates acyl-CoA bound fatty acid substrates in the cytosolic acyl-CoA pool as opposed to desaturating phosphatidylcholine bound fatty acid substrates in the endoplasmic reticulum-associated phospholipid pool.
262 No definition of “substrate specificity” is provided in the application as filed. But in my view the person skilled in the art would take it to refer to the ability of an enzyme to recognise and bind to particular substrates such as a fatty acid in a manner relevant to the context of catalytic conversion and in the context of the type of biosynthetic pathway under discussion.
263 Now fatty acid desaturases will have different abilities to interact with different fatty acid substrates, that is, fatty acids with different chain lengths or C=C double bonds at different positions. Accordingly, they will have different substrate specificities.
264 An enzyme categorised as a Δ6-desaturase will at least have a substrate specificity for one of, the ω6 fatty acid substrate, LA or the ω3 fatty acid substrate, ALA, because it catalyses Δ6-desaturation of these substrates to produce SDA and GLA respectively. But the same enzyme may also act on shorter or longer fatty acid substrates in addition to ALA and LA, or may act on fatty acids having C=C double bonds in different positions. Further, the enzyme may also introduce a C=C double bond at a different position in a fatty acid substrate, for example at the Δ8 position or at the Δ5 position, in addition to the Δ6 position. For this reason, an enzyme described as a Δ6-desaturase may have substrate specificity for a number of different fatty acid substrates.
265 Now two Δ6-desaturases that both act on only ALA and LA would have the same substrate specificity. But if the second enzyme acted on ALA and LA and also acted on ETA, that is, it introduced a C=C double bond at the fifth carbon atom from the terminal carboxyl end therefore having Δ5-desaturase activity too, it could be said to have a different substrate specificity. The second enzyme would have a substrate specificity for three substrates (ALA, LA and ETA), whereas the first enzyme would only have a substrate specificity for two substrates (ALA and LA).
266 Now Dr Stalker in his affidavit said:
… I read ‘a CoA-dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID No: 14’, to be simply describing an enzyme that functions as a Δ6-desaturase and, thus has an affinity for ALA and/or LA. I do not interpret this as meaning that the Δ6-desaturase has to have identical substrate specificity of the Δ6-desaturase of SEQ ID NO: 14. … That is, in my view, the substrate specificity of a Δ6-desaturase encompasses only substrates which undergo Δ6-desaturation being LA and/or ALA.
267 Further, Dr Stalker stated that “in the context of [the schematic reproduced earlier in my reasons], the substrate specificity for an enzyme functioning as a Δ6-desaturase acts on LA or ALA or both; any other specificity or side chain reaction is not relevant to that Δ6-desaturase activity”. Dr Stalker said, in effect, that the overall substrate specificity of the Δ6-desaturase show in SEQ ID NO: 14 is irrelevant. As I understood his evidence, Dr Stalker therefore appeared to limit the meaning of “substrate specificity” to a Δ6-desaturase’s primary catalytic activity only.
268 Further, in Dr Stalker’s opinion set out in the joint expert report, “a CoA dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14” means that any enzyme that carries out a Δ6 desaturase catalytic reaction is specified in the first bullet point of the bridging paragraph that it have “the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14”, ie, that the enzyme must act on a substrate that can desaturate at the 6-carbon position of a fatty acid substrate molecule. So, in essence, such an enzyme can be categorised as a Δ6 desaturase by having substrate specificity for at least one of the ω3 or ω6 fatty acids; and so ALA or LA or both as substrates. Further, he said that there is no provision in the bullet point for a CoA dependent Δ6 desaturase having “the same” or “identical” substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14.
269 Further, Dr Stalker made reference to Dr Singh’s Table that I have set out earlier. According to Dr Stalker, in the examples of Δ6 desaturases from organisms displayed in that Table, O. tauri, O. RCC809 and M. pusilia Δ6 desaturase enzymes all have the ability to covert ALA to SDA, so in essence have “the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14” that is described in the application as filed. The O. lucimarinus enzyme also has that capability. He said that whether these enzymes can convert LA to GLA, or not produce any detectable level of GLA (the O. RCC809 enzyme), or catalyze other side reactions (the Δ5 or Δ8 reactions on ETA and ETrA substrates by the O. tauri enzyme) is not germane to the fact that all Δ6 desaturases described in the Table have the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14.
270 I do not accept BASF’s position or Dr Stalker’s analysis on this aspect.
271 Dr Singh did not read the substrate specificity feature as only referring to the substrates that undergo Δ6-desaturation. This is because the meaning of “substrate specificity” is not limited to the primary catalytic activity of the enzyme. In his view, the term “substrate specificity” refers to the ability of an enzyme to recognise and bind to particular substrates, and to not bind to or act upon other related molecules within the context relevant to catalytic activity. That is, the phrase “the substrate specificity” is not limited to the substrates the enzyme binds to as part of its primary function. Rather, “substrate specificity” is understood in the art to refer to all substrates a particular enzyme can bind to and to potentially catalyse some type of reaction. He defined an enzyme’s specificity as the ability to discriminate between a substrate and a competing molecule. Moreover, he said that specificity was not limited to the enzyme’s primary catalytic activity. I accept that evidence providing that it is understood in the context of the biosynthesis contemplated in the application as filed, that is, relevant potential substrates.
272 Dr Singh said in the joint expert report that an enzyme can be categorised as a Δ6 desaturase by having a substrate specificity for at least one of the ω3 or ω6 fatty acids, namely, the ALA and/or LA substrates via insertion of a Δ6 double bond to produce the SDA and/or GLA products respectively. But the same enzyme may also act on shorter or longer fatty acid substrates in addition to ALA or LA, or other fatty acids having a C=C double bond at different positions of the respective fatty acid molecule. Further, a Δ6 desaturase may also introduce a C=C double bond at a different position, in addition to the Δ6 position. For this reason and as I have said, an enzyme described as a Δ6 desaturase may have substrate specificity for other different fatty acid substrates. Two Δ6 desaturases that act only on ALA and LAs substrates would have the same (identical) substrate specificity. But as I have said, assume that the second enzyme of the two Δ6 desaturases can also act on the substrate ETA to introduce a C=C double bond at the 5th carbon atom from the carboxyl end (a Δ5 desaturase activity). It would have a different substrate specificity. The second enzyme would have a substrate specificity for three substrates (ALA, LA and also ETA), whereas the first enzyme would only have a substrate specificity for two substrates (ALA and LA). The Δ6 desaturase from O. tauri has activity on ALA, LA as well as ETA while the Δ6 desaturase from O. lucimarinus (SEQ ID NO: 14) does not have activity on ETA. Hence these two enzymes do not have the same substrate specificities. Further, some enzymes can catalyse more than one type of reaction. This is exemplified by O. tauri and Micromonas Δ6 desaturases. Further, O. tauri Δ6 desaturase can also insert a Δ6 double bond in the monounsaturated fatty acid oleic acid to produce isolinolenic acid. Further, unlike O. tauri and Micromonas Δ6 desaturases, SEQ ID NO: 14 has no detectable specificity for the substrates ETA and ETrA.
273 Now the bridging paragraph refers to the enzyme as being a CoA-dependent Δ6-desaturase because that is the enzyme’s primary catalytic activity. But the CoA-dependent Δ6- desaturase may be able to undertake other catalytic activities, for example a Δ5-desaturation at the Δ5 position. If so, it would still be referred to as a Δ6-desaturase because that activity is the enzyme’s primary function. However, according to Dr Singh, the reference to “the substrate specificity” in the bridging paragraph was not limited to the substrates the enzyme binds to as part of its primary catalytic activity. I must say that I agree with Dr Singh.
274 Further, Dr Singh also considered that Dr Stalker’s reading of the bridging paragraph gave the words “having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14” no work to do. Dr Stalker read the feature as referring to any CoA-dependent Δ6-desaturase rather than “a CoA-dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14”. Accordingly, Dr Stalker’s reading gives no work to do to the qualifier.
275 Further, Dr Singh disagreed with Dr Stalker’s interpretation that the phrase “the substrate specificity” does not mean the same relevant substrate specificity. This is because the substrate specificity feature specifically refers to the CoA-dependent Δ6-desaturase “having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14” (my emphasis). Dr Singh understood the word “the” to denote a specific or particular effect. For this reason, he read the substrate specificity feature as saying that the invention is directed to a CoA-dependent Δ6-desaturase having “the same” or “the specific” or “the identical” substrate specificity of “the Δ6 desaturase shown in SEQ ID No:14”. And this therefore requires a comparison with the specific substrates that the Δ6-desaturase shown in SEQ ID NO: 14 binds to, being the substrates that the enzyme binds to as part of its primary activity (being a Δ6 desaturase) and any substrates that enzyme binds to as part of any side reactions, related reactions or bifunctional reactions, but of course relevant to the biosynthetic pathway in question.
276 Further, Dr Singh said that although a Δ6-desaturase will predominantly desaturate LA or ALA to produce GLA or SDA respectively, it has also been shown to bind to oleic acid (OA) to produce isolinoleic acid through desaturation at the sixth carbon. For example, a recent paper detailed a study showing that the Δ6-desaturase from Phytophthora citrophthora transformed in Perilla seeds bound to OA, LA and ALA to produce isolinoleic acid, GLA and SDA respectively (Lee et al, High accumulation of γ-linolenic acid and Stearidonic acid in transgenic Perilla (Perilla frutescens var. frutescens) seeds, BMC Plant Biology (2019) 19:120). Further, the O. tauri Δ6-desaturase has also been reported to have Δ6-desaturase activity on OA to produce isolinoleic acid; see Domergue that I cited earlier. Accordingly, Dr Singh did not read the substrate specificity feature as encompassing only the substrates LA or ALA.
277 In my view, the expression “the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14” (my emphasis) serves to describe the specificity which is unique or specific to that particular polypeptide relevant of course to the biosynthetic context one is considering.
278 The requirement that the CoA-dependent ∆6-desaturase have the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14 defines and limits the substrate specificity of the CoA ∆6-desaturase described and claimed to those which possess the particular specificity of the ∆6-desaturase shown in SEQ ID NO: 14.
279 Now a ∆6-desaturase will at least have a substrate specificity for ALA and/or LA. This is its primary catalytic activity. But a ∆6-desaturase may also have substrate specificity for different fatty acid substrates.
280 Dr Stalker wrongly equated the primary catalytic activity of any ∆6-desaturase (i.e. acting on ALA and/or LA) with the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14. Primary catalytic activity is not the same as “the substrate specificity”.
281 Further, Dr Stalker’s reading would render the words “having the substrate specificity of the ∆6-desaturase shown in SEQ ID NO: 14” redundant. This is because a “CoA-dependent Δ6 desaturase”, without more, already describes any “enzyme that functions as a ∆6-desaturase and thus has an affinity for ALA and/or LA.”
282 Further, Dr Stalker’s interpretation ignores the qualifying nature of the word “the”.
283 Further, although a ∆6-desaturase will predominantly desaturate LA and/or ALA, it may also have ∆6-desaturase activity on OA to produce isolinoleic acid by desaturation at the sixth carbon. Accordingly, even if this expression referred only to the ∆6-desaturase activity of an enzyme, it would not be limited to the substrates LA and ALA.
284 In the context of the application as filed, in my view “the substrate specificity” refers to all of the fatty acid substrates within the LC-PUFA pathway that the enzyme binds to.
285 As to the CoA-dependent Δ6-desaturase having the relevant substrate specificity, it must have all of the fatty acid substrates of the Δ6-desaturase shown in SEQ ID No: 14 within the LC-PUFA pathway. Examples of such substrates for Δ6-desaturases within the LC-PUFA pathway include LA, ALA, OA. But it is not required to have those substrates of the Δ6-desaturase shown in SEQ ID No: 14 that do not relevantly form part of the LC-PUFA pathway (if any); such substrates would be understood by the skilled addressee to be extraneous to the relevant context.
286 Now I agree with CSIRO that it is not for it to advance a practical explanation as to why the patent applicant chose to limit its description of the invention in this manner. The specification of a patent is a document in words of the patentee’s own choosing, usually drafted with highly skilled and expensive advice. A patentee may have good reason for introducing a limitation into a claim, including to avoid arguments in relation to prior art. For example, I note that Domergue discloses a Δ6-desaturase that binds to and converts four substrates in the LC-PUFA pathway, including ALA and LA. It may be speculated that if the first bullet point of the bridging paragraph did no more than describe a Δ6-desaturase that binds to and converts ALA and LA, the invention may have been vulnerable to attack on the ground of lack of novelty.
287 BASF’s construction involves impermissibly ignoring the limitation the patentee chose to introduce into the claim, that is, “having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14”.
288 Further, BASF’s construction involves adding a gloss to the words of the disclosure such as core or primary substrate specificity.
289 Further, on BASF’s construction, a Δ6-desaturase acting on two substrates within the LC-PUFA pathway would have the same substrate specificity as a Δ6-desaturase acting on four substrates in that pathway because they have two common substrates. But there is no cogent reason that would support such a conclusion.
290 In summary, I reject BASF’s construction involving “substrate specificity”. This conclusion is also sufficient to dispose of BASF’s tertiary argument.
(b) Preference for conversion of ALA compared to LA
291 In addition to the CoA-dependent Δ6-desaturase having the same substrate specificity (in context) as the Δ6-desaturase shown in SEQ ID NO: 14, the application as filed also states that the CoA-dependent Δ6-desaturase is to have a preference for conversion of the ω3-fatty acid, ALA compared to the ω6-fatty acid, LA. That is, it operates preferentially in the ω3 pathway, rather than the ω6 pathway shown in the schematic that I set out earlier in my reasons.
292 Now no definition of “preference” is provided in the application as filed. The only two references to “preference” in the application as filed are on page 7 line 1 and claim 2.
293 But it is not in dispute that the term “preference” in this context refers to a quantitative comparison of the rate of conversion of ALA to SDA relative to the conversion of LA to GLA. Preference refers to greater desaturase activity on ALA relative to LA. On this basis, in any relevant study involving the claimed CoA-dependent Δ6-desaturase, it would be expected that the resulting product would contain a higher amount of SDA than GLA, relative to the amount of ALA and LA added in the study, as a result of the claimed CoA-dependent Δ6-desaturase’s preference to convert ALA into SDA.
294 Now I should say at this point that the concepts of conversion preference and conversion rate should not be confused. The former deals with a comparison of rates. Let me return to the application as filed.
295 The application as filed describes a CoA-dependent Δ6-desaturase having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14 and also having a preference for conversion of ALA compared to LA. This is because of the use of the conjugative “and” at page 6 line 34 which refers to ALA conversion preference as being in addition to the requirement of identical substrate specificity. This conclusion is further supported by the reference to “the above” CoA-dependent Δ6-desaturase at page 7 line 1, which defines the feature of ALA conversion preference by reference to, and combined with, the previously mentioned feature of substrate specificity.
296 Now in the context of the bridging paragraph, the statement disclosed in the first bullet point can be independent of the statement disclosed in the second bullet point. So, the statement disclosed in the first bullet point, ie, the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14 does not need to include the ALA preference feature disclosed in the second bullet point. But the words “the above” entail that the statement disclosed in the second bullet point needs to be read in context with the statement in the first bullet point.
297 The experts were asked to consider the application as filed, and explain whether they considered that it disclosed a CoA dependent Δ6-desaturase that preferentially converts ALA compared to LA. They gave the following evidence. Preference in context is a qualitative comparison of the rate of conversion of ALA to SDA relative to the rate of conversion of LA to GLA. On this basis, in any expression test involving the claimed CoA-dependent Δ6 desaturase, they would expect that the enzymatic reaction would produce a higher amount of SDA than GLA. Now in the application as filed, a Δ6 desaturase that has a preference for ALA as a substrate does not define the degree of preference for ALA as compared to LA as a substrate. And there is no specific data in the application as filed showing the degree of preference for the Δ6 desaturase shown in SEQ ID NO: 14 to convert ALA over LA. Further, there is no data in the application as filed showing preference for ALA as a substrate for any of the other O. lucimarinus sequences disclosed. The only mention of the words “preference for ALA” in the application as filed occurs in the bridging paragraph and claim 2. There is a reference to “[e]specially preferably” on page 7 line 13, and in that context the page 8 paragraph refers to Δ6 desaturase activity. But those descriptions do not broaden the scope of the disclosure. The page 8 paragraph is to be read in the context of the subject of the invention of the application as filed i.e. a CoA-dependent Δ6 desaturase having the substrate specificity of SEQ ID NO: 14 (claim 1) and that this CoA-dependent Δ6 desaturase may also have a substrate preference for ALA over LA (claim 2).
298 Further, claim 1 of the application as filed claims “a CoA-dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14”. That is claim 1 claims only the feature of substrate specificity.
299 Claim 2 of the application as filed claims “the CoA-dependent Δ6 desaturase according to claim 1, wherein the desaturase has a preference for conversion of [ALA] compared to [LA]”. That is, claim 2 claims both of the features of substrate specificity and ALA conversion preference.
300 Accordingly, the claims of the application as filed are consistent with the interpretation that the preference for conversion of ALA compared to LA is a feature of a CoA-dependent Δ6-desaturase that also has the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14.
301 The experts were asked to consider whether the application as filed disclosed a CoA-dependent Δ6-desaturase that does not have the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14 but which preferentially converts ALA compared to LA. They agreed that there was no such express disclosure.
302 In summary, the application as filed discloses a CoA-dependent Δ6-desaturase that has the ALA conversion preference which also has the same substrate specificity as the CoA-dependent Δ6-desaturase shown in SEQ ID NO: 14. There is no disclosure in the application as filed of a CoA-dependent Δ6-desaturase that has the ALA conversion preference without such CoA-dependent Δ6-desaturase also having the same substrate specificity of the CoA-dependent Δ6-desaturase shown in SEQ ID NO: 14.
303 The experts were also asked to consider whether a CoA-dependent Δ6-desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 and which preferentially converts ALA compared to LA was disclosed in the application as filed.
304 Dr Singh’s opinion was no. The application as filed is limited to a CoA dependent Δ6 desaturase both having the substrate specificity shown in SEQ ID NO: 14 and having a preference for ALA over LA as a substrate. There are 2 scenarios that he said were worth considering here.
305 First, it is possible that a Δ6 desaturase having at least 75% homology at the DNA level to the Δ6 desaturase shown in SEQ ID NO: 14 also has the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14.
306 Second, it is also possible that a Δ6 desaturase, where the DNA coding region has at least 75% homology at the DNA level to the sequence that codes for the polypeptide shown in SEQ ID NO: 14, exhibits a more relaxed substrate specificity than the Δ6 desaturase shown in SEQ ID NO: 14. Examples of Δ6 desaturases cited in Dr Singh’s Table that I have reproduced earlier illustrate this scenario. For example, the O. tauri Δ6 desaturase has 79% nucleotide homology to SEQ ID NO: 13 (coding for the polypeptide shown in SEQ ID NO: 14) of the application as filed. The O. tauri Δ6 desaturase can also convert the substrate ETA to EPA, whereas the polypeptide coded for by SEQ ID NO: 13 does not perform that function. It is noteworthy that conversion of ETA to EPA is in the context of the entire PUFA pathway shown in the biosynthetic pathway schematic that I have set out earlier. In fact, a single amino acid alteration could alter the function and thereby change the substrate specificity of an enzyme. So, it is also possible that two enzymes can have different substrate specificities and have a 75% homology to each other at the DNA level. Dr Singh said that due to the existence of this scenario, a 75% comparative homology at the DNA level for a Δ6 desaturase cannot be taken to mean that such a Δ6 desaturase has the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14 in the application as filed. I must say that I had no reason to doubt such evidence.
307 One can contrast all of this with the proposed amendments.
308 The amendments, by seeking to insert new claims 2 and 7 into the application as accepted, would claim a CoA-dependent Δ6-desaturase:
(a) having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 (see claims 1 and 5 (now claim 6) of the application as accepted); and
(b) that preferentially converts ALA compared to LA (proposed new claim 2, when read together with claim 1, of the application as accepted).
309 But the second matter is new matter that was not originally disclosed in the application as filed. This is because this feature of a CoA dependent Δ6-desaturase that preferentially converts ALA compared to LA was only ever disclosed in the context of a CoA-dependent Δ6-desaturase that has the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14.
310 By contrast, a CoA dependent Δ6-desaturase having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14 and the ALA conversion preference as claimed in the application as filed is not the same as a CoA-dependent Δ6-desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 and having the ALA conversion preference (as claimed in new claims 2 and 7).
311 So new claims 2 and 7 would be seeking to claim a CoA-dependent Δ6-desaturase that was not disclosed in the application as filed. This is because the application as filed was limited to a CoA-dependent Δ6-desaturase that has both the identical substrate specificity and ALA conversion preference features, whereas new claim 2 is seeking to claim a CoA-dependent Δ6-desaturase that has the ALA conversion preference only, together with having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14, which is not the same as identical substrate specificity.
312 Let me say something about the amendment which seeks to insert the following description into the body of the specification of the application as accepted (at page 7b):
According to an embodiment of the abovementioned process and use, the CoA-dependent desaturase preferentially converts [ALA] compared to [LA].
313 This amendment is seeking to describe a CoA-dependent Δ6-desaturase having this preference for ALA, but where the CoA-dependent Δ6-desaturase has at least a 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 (as opposed to having the substrate specificity of SEQ ID NO: 14).
314 But given that substrate specificity and at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 are different, this amendment, by seeking to insert this new paragraph into page 7b of the application as accepted, would be seeking to add new material that was not disclosed in the application as filed in terms of amounting to an impermissible intermediate generalisation.
(c) Page 8 paragraph
315 Page 7, line 20 states, “Therefore, polynucleotides according to the invention are in particular:...”. Page 7 line 21 to page 8 to line 27 describe polynucleotides that are preferably expressed in the host cell or transgenic non-human organism encoding the suite of enzymes involved in the production of fatty acids, specifically DHA, as per the pathway shown in the figure for the schematic representation of the conversion of LA or ALA. Specific polynucleotides described include SEQ ID NO: 1, 3, 5, 6, 7, 9, 11, 13, 15 or 16 encoding a suite of enzymes, including a Δ4-desaturase, a Δ5-desaturase, a Δ5-elongase, a Δ6-desaturase a Δ6-elongase and a Δ12 desaturase. Although elongases and desaturases other than Δ6-desaturases are referred to in this paragraph and elsewhere in the specification, the invention as claimed in both the application as filed and the application as accepted is limited to certain Δ6-desaturases.
316 In relation to the Δ6-desaturase, the page 8 paragraph is “therefore” an example of “the invention”. In my view this entails that the polynucleotides in part (iii) of the page 8 paragraph must have the required substrate specificity coupled with the ALA conversion preference as described in the bridging paragraph.
317 The skilled addressee does not read the page 8 paragraph in isolation. The page 8 paragraph discloses the polynucleotides “according to the invention” (application as filed, page 7, line 20). The page 8 paragraph does not broaden the scope of the invention disclosed. It is to be read in the context of the bridging paragraph.
318 Now Dr Stalker gave evidence that parts (iii) and (iv) of the page 8 paragraph provide a broader range of polynucleotides than (i) and (ii), provided that the Δ6-desaturase activity is preserved. But polynucleotides within parts (iii) and (iv) must also preserve the required substrate specificity and ALA conversion preference features as disclosed in the bridging paragraph.
319 The descriptions which follow on pages 9 to 11 and page 30 lines 1 to 2 and 25 to 28 of the application as filed are subject to the same limitation.
320 Further, in relation to a Δ6-desaturase which has the ALA conversion preference, the application as filed only discloses and claims a Δ6-desaturase as having this feature, in the context of the Δ6-desaturase also having “the substrate specificity” feature of the Δ6- desaturase shown in SEQ ID NO: 14. That is, the application as filed does not disclose the ALA conversion preference independently of the required substrate specificity feature.
321 Now although Dr Stalker acknowledged the conjugative “and” in the bridging paragraph at the end of the first bullet point, insufficient attention was given to the reference to “the above” at page 7 line 1. But the reference to “the above” defines the ALA conversion preference feature by reference to, and combined with, the previously mentioned feature of the required substrate specificity.
322 Now BASF contends that read in isolation, there is a disclosure in the page 8 paragraph of a polynucleotide coding for a Δ6 desaturase having the amino acid sequence disclosed as SEQ ID NO: 14 and of certain polynucleotides within specified ranges of sequence homology. Further, it contends that the second bullet point of the bridging paragraph discloses that the SEQ ID NO: 14 polypeptide preferentially converts ALA compared to LA.
323 Therefore, the skilled addressee understands from the application as filed that:
(a) Δ6 desaturases will act in LC-PUFA biosynthetic pathways; and
(b) as a subset of the above, Δ6 desaturases will preferentially convert ALA to LA; and
(c) polynucleotides having the requisite homology (75% identity) will likely produce polypeptides that preserve those biological functions.
324 But BASF impermissibly seeks to strip the conversion preference (feature B) from the context in which it is disclosed in the bridging paragraph, by attempting to describe it as a property of SEQ ID NO: 14 and therefore combine feature B with the required sequence homology (feature C). But feature C is not coterminous with the required substrate specificity (feature A).
325 Further, BASF’s argument proceeds on a flawed foundation.
326 The conversion preference was to be applied to the CoA-dependent Δ6 desaturase having the substrate specificity of the Δ6 desaturase shown in SEQ ID NO: 14. This feature was not directly referable to the Δ6 desaturase shown in SEQ ID NO: 14 itself.
327 The bridging paragraph does not disclose the conversion preference of SEQ ID NO: 14. The skilled addressee would not understand from the second bullet point of the bridging paragraph or any part of the application as filed that one of the properties of SEQ ID NO: 14 itself is that it will preferentially convert ALA compared to LA.
328 First, there is no data in the application as filed to support that proposition.
329 Second, the bridging paragraph discloses a class of enzymes that answer the description of “a CoA-dependent Δ6-desaturase having the substrate specificity of the Δ6-desaturase shown in SEQ ID NO: 14.” Accordingly, the disclosure of the class of enzymes described in the first bullet point is not a disclosure of the conversion preference for SEQ ID NO: 14. That is, the substrate specificity of the class of enzymes described in the first bullet point is not determinative of the conversion preference of those enzymes.
330 Third, such a class is narrowed in the second bullet point. But it does not disclose a property of SEQ ID NO: 14.
331 Fourth, the difference in language between the first bullet point and the second bullet point is significant. It does not describe the conversion preference by reference to the Δ6-desaturase shown in SEQ ID NO: 14.
332 In summary, in my view the page 8 paragraph does not disclose the invention the subject of the proposed amendment. The specific processes and uses BASF now seeks to disclose and claim with respect to a CoA dependent Δ6-desaturase with feature C combined with feature B is, at best, a “subset” of the page 8 paragraph considered in isolation. But in my view the skilled addressee would not have read the page 8 paragraph in isolation in the application as filed. And nor would they have read the second bullet point of the bridging paragraph in isolation. In my view, a person skilled in the art would not so read the page 8 paragraph out of context as Dr Stalker has done.
333 So, the page 8 paragraph of the application as filed does not disclose a CoA-dependent Δ6-desaturase:
(a) having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 (proposed new claim 6); and
(b) that preferentially converts ALA compared to LA (proposed new claim 2, when read together with claim 1, of the application as accepted).
334 Accordingly, BASF’s primary argument should be rejected.
(d) Other matters
335 Is anything left of the secondary argument? I do not think so. In my view one cannot diminish the significance of the bridging paragraph as BASF has sought to do. Now I accept that the bridging paragraph discloses the conversion preference. But as I say, that was in a specific and in substance cumulative context.
336 In summary, BASF by the amendment seeks to take the conversion preference feature well outside the context of the application as filed. In the application as filed it was limited to a CoA dependent Δ6 desaturase that also had the substrate specificity of SEQ ID NO: 14. So, it was dealing with such a desaturase that had both features, not the conversion preference feature alone. Further, the preferments were to be understood in the context of the bridging paragraph. Further, the page 8 paragraph did not describe a broader invention. Rather, the polynucleotides were those which coded for the polypeptides of the invention described in the bridging paragraph.
337 So, when one considers the application as filed read as a whole through the eyes of the skilled addressee, it relevantly disclosed the invention as:
(a) a CoA-dependent Δ6 desaturase with the relevant substrate specificity; and
(b) a CoA-dependent Δ6 desaturase having both the relevant substrate specificity and the conversion preference.
338 Contrastingly, the proposed amendments take the conversion preference feature (feature B) and add it to the feature of a CoA-dependent Δ6 desaturase having at least 75% identity to a nucleotide sequence which codes for a polypeptide as shown in SEQ ID NO: 14 (feature C). But this was not disclosed in the application as filed. To put it bluntly, the relevant substrate specificity (feature A) and feature C are not the same.
339 Moreover, even if the relevant disclosure in the application as filed is not limited to the invention, the amendment is still not allowable. Even though the page 8 paragraph or other stray references may include or cover the matters in the application as proposed to be amended, it does not disclose those matters for the purpose of s 102(1). It does not disclose the specific processes and uses it now seeks to disclose and claim with respect to feature C combined with feature B.
340 If I have not already made it clear, you cannot just take the page 8 paragraph as free-standing and add to it feature B. To do so you are decontextualising both. Both are to be read in the context of the bridging paragraph. And nor is it permissible to just take the page 8 paragraph as free-standing and to say that the page 8 paragraph together with feature B is just a sub-set. Again that is impermissibly decontextualising.
341 Accordingly, the proposed amendments constitute an impermissible intermediate generalisation. Let me conclude by making the following other points to the extent that I have not already done so.
342 First, BASF has said that features A and B from the bridging paragraph are not combined in the sense of a combination of physical integers. They each describe properties of an individual polypeptide. Now this may be accepted. But in my view, so to accept does not deny the force of CSIRO’s point concerning intermediate generalisation.
343 Second, BASF also says that features A and B do not co-operate to achieve some unitary effect or outcome. But even if this be so, that does not deny the proposition that the re-addition of feature B amounts to an impermissible intermediate generalisation.
344 Third, BASF has also sought to place some emphasis on the passage in the application as filed which begins “Especially preferably…” (p 7 line 13). But in the context of Δ6 desaturases, this is not broadening from the bridging paragraph. And in terms it is not referring to the conversion preference in the second bullet point of the bridging paragraph.
345 Fourth, let me say something further concerning claims. BASF says that although claims of the complete specification as filed can form part of the disclosure, they do not limit the scope of the disclosure. I agree. But this does not really take BASF anywhere.
346 Fifth, BASF says that the amendments are narrowing rather than broadening. As I have said, seen in the context of the application as accepted, they are narrowing. But seen in the context of a comparison with the application as filed, that is not a satisfactory description. In any event, I prefer not to use a narrowing/broadening discriminant. That is not the language of s 102(1).
347 Finally, let me make one other observation. I have not engaged in any discussion as to whether the proposed amendments would have also been disallowed under the old version of s 102(1). I prefer to avoid any retrospective hypothetical.
348 The appeal is allowed. The proposed amendments are impermissible. I will make the necessary orders.
I certify that the preceding three hundred and forty eight (348) numbered paragraphs are a true copy of the Reasons for Judgment herein of the Honourable Justice Beach. |
Associate:
Schedule